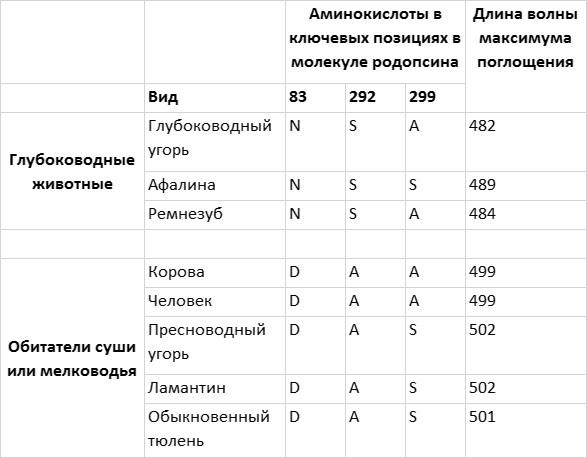
Рис. 4.6. Настройка родопсина зависит от глубины, на которой обитают морские животные. Глубоководные рыбы и китообразные имеют родопсин, максимум поглощения которого сдвинут в синюю область спектра по сравнению с родопсинами животных, обитающих на суше или на мелководье. У животных, обитающих на одинаковой глубине, часто в ключевых позициях в последовательности родопсина стоят одни и те же аминокислоты. Рисунок Джейми Кэрролл.
Связь «синего сдвига» родопсина с адаптацией к жизни в морских глубинах подкрепляется данными анализа родопсинов глубоководных и пресноводных угрей. Родопсин угря, обитающего на большой глубине, характеризуется сдвигом максимума поглощения в синюю область и в ключевых положениях имеет точно те же аминокислотные остатки, что и родопсин ремнезуба. Родопсин пресноводного угря имеет максимум поглощения, близкий к таковому у наземных млекопитающих, а в ключевых позициях в его последовательности находятся те же аминокислотные остатки, что и у ламантина и тюленя — двух обитающих на мелководье млекопитающих с типичным вариантом родопсина.
Эта поразительная связь между «синим сдвигом» родопсина и жизнью на большой глубине полна глубокого смысла. Резонно предположить, что естественный отбор способствовал настройке родопсина в соответствии со средой обитания. Однако существует еще более важная причина подозревать влияние естественного отбора, и она становится ясна при рассмотрении эволюционных связей между видами животных, перечисленных на рис. 4.6. Дельфины и ремнезубы относятся к отряду китообразных — млекопитающих, которые произошли от сухопутного предка, вернувшегося в водную стихию. Это может показаться удивительным, но ближайшими современными родственниками китообразных являются гиппопотамы, олени, коровы, свиньи и верблюды. Это следует из результатов анализа последовательностей SINE и LINE, а также других последовательностей ДНК. Поскольку родопсины наземных родственников китообразных настроены на длину волны 500 нм, можно с полным основанием заключить, что родопсин дельфинов и китов изменился после того, как их эволюционная ветвь отделилась от ветви других млекопитающих.
Однако угри — это рыбы, и их эволюционная ветвь отделилась от ветвей других позвоночных несколько сотен миллионов лет назад. Это означает, что одинаковые по своей сути различия между глубоководными и пресноводными угрями и морскими и наземными млекопитающими возникли независимо. Если в одинаковых позициях в каком-то белке у двух или нескольких видов животных при адаптации к определенным условиям появляются одни и те же аминокислотные остатки (как у китообразных и глубоководного угря в нашем примере), это говорит о работе естественного отбора. Замечу, что пример эволюции родопсина — лишь один из множества случаев, когда эволюция воспроизводит саму себя (мы подробнее поговорим об этом в шестой главе).
Теперь перейдем от красного, зеленого и синего к невидимым для нас цветам — и к целому миру взаимоотношений между животными, связанных с ультрафиолетовым зрением.
За пределами радуги
Примерно через 12 лет после выхода в свет книги «О происхождении видов» Дарвин опубликовал «Происхождение человека и половой отбор». Это был первый труд Дарвина, посвященный эволюции человека, но, возможно, еще важнее то, что Дарвин впервые обратил внимание на различную роль полов в эволюции признаков; он называл этот процесс «половым отбором». И сам половой отбор, и развитая Дарвином теория полового отбора известны широкой публике гораздо хуже, чем его теория естественного отбора, однако биологи считают половой отбор одним из наиболее важных и интересных механизмов в эволюции животных. Половой отбор — это арена, на которой «наилучшая приспособленность» оказывается напрямую связана с успешностью спаривания.
Дарвин был увлечен разнообразием птичьей окраски — он посвятил много времени изучению этого удивительного явления и оставил многостраничные описания роскошного оперения самых разных видов птиц. Особенно его интересовало, каким образом предпочтения самок могут привести к эволюции причудливого оперения самцов, такого как павлиний хвост. Еще со времен Дарвина птицы остаются излюбленным объектом биологов, изучающих половой отбор. Но до недавних пор подобные исследования имели один и тот же изъян: окраску птиц оценивали люди. Я уже говорил о том, что мы видим мир совсем не таким, каким его видят другие млекопитающие, и не таким, каким его или друг друга видят птицы. Многие птицы способны видеть в ультрафиолетовом свете и различать цвета, не существующие для нас, и эта способность играет важную роль в спаривании, поиске пищи и даже вскармливании птенцов.
У многих птиц опсины настроены на восприятие ультрафиолетового света, и на их оперении есть отметины, отражающие свет в ультрафиолетовой части спектра (рис. D-K).
Длина волны ультрафиолетового света меньше 400 нм, то есть меньше длины волны фиолетового света, и этот свет регистрируется коротковолновым опсином. Человеческий опсин SWS настроен на длину волны 417 нм. У некоторых видов птиц опсины SWS настроены на длину волны около 370 нм, поэтому эти птицы видят в ультрафиолетовом свете. Другие птицы имеют коротковолновые опсины, настроенные на фиолетовую область спектра, с максимумом поглощения около 405 нм, и эти птицы, как и люди, не могут воспринимать ультрафиолетовый свет. Лабораторные исследования помогли установить, какие изменения опсинов SWS отвечают за чувствительность птиц к ультрафиолету.
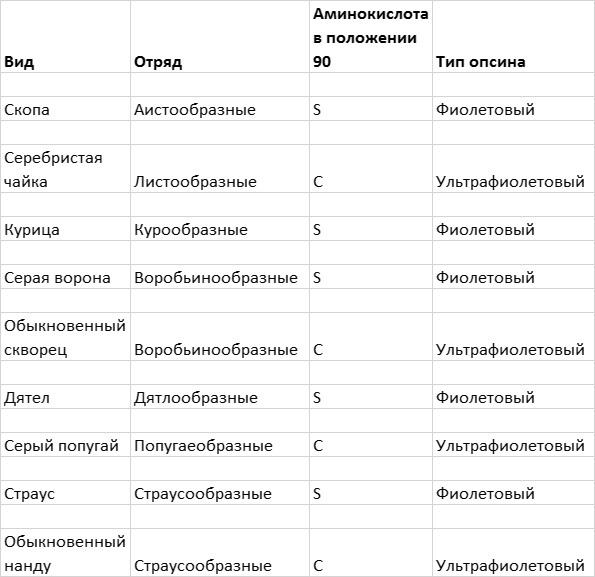
Одна специфическая позиция, а именно позиция 90 в коротковолновом опсине, напрямую связана со способностью птиц воспринимать фиолетовый или ультрафиолетовый свет. Те птицы, у которых в этом положении находится аминокислота серин, видят в фиолетовой области спектра, а те, у которых в этом месте располагается цистеин, видят в ультрафиолетовом диапазоне (рис. 4.7).
Рис. 4.7. Эволюция способности птиц к восприятию ультрафиолетового света. Восприятие птицами фиолетового или ультрафиолетового света в значительной степени зависит от того, какая аминокислота — серин (S) или цистеин (C) — находится в позиции 90 в молекуле коротковолнового опсина. Замена аминокислоты в этой позиции в разных отрядах птиц происходила не менее четырех раз. Рисунок Джейми Кэрролл.
Седзо Йокояма, один из ведущих специалистов в области цветового зрения, и его коллеги из Университета Эмори (Джорджия) напрямую показали, что замена серина на цистеин делает пигмент, чувствительный к фиолетовому свету, чувствительным к ультрафиолетовому свету, а замена цистеина на серин в пигменте, чувствительном к ультрафиолету, делает его восприимчивым к фиолетовому свету. Разница в одной-единственной аминокислоте обеспечивает сдвиг максимума поглощения на 35–38 нм, что чрезвычайно много. Эти исследования показывают, что одна замена способна изменить функцию опсина, и следовательно, эволюция опсина, чувствительного к фиолетовому или ультрафиолетовому свету, может быть относительно простым, одностадийным процессом.
Птицы, обладающие способностью видеть в ультрафиолетовом свете, относятся к девяти семействам из четырех разных отрядов. На основании их родственных связей можно сделать вывод, что у птиц способность видеть в ультрафиолетовом свете возникала в ходе эволюции независимым образом не менее четырех раз. Во всех отрядах, к которым относятся птицы, способные воспринимать ультрафиолет, есть также птицы с фиолетовым типом SWS-опсина. Это означает, что мутация, заменившая серин на цистеин в молекуле фиолетового пигмента, происходила в ходе эволюции несколько раз.
В ситуации с ультрафиолетовым зрением эволюция вновь повторяет саму себя. Это убедительное доказательство того, что на гены опсина оказал воздействие отбор. Однако в данном случае это мог быть половой отбор, поскольку многочисленные исследования показывают, что на выбор партнера у птиц, способных видеть в ультрафиолетовой области спектра, влияет окраска, различимая только в ультрафиолетовом диапазоне (рисунки D-K цветной вкладки).