Рис. 3. Расположение материков в начале кайнозойской эры
Первые обезьяны
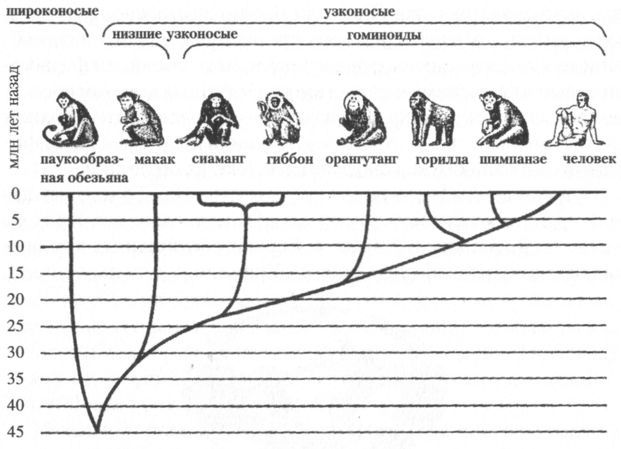
В раннем эоцене (54–45 млн лет назад) в пределах отряда приматов выделяется уже множество семейств, родов и видов, среди которых есть и предки современных лемуров и долгопятов. Обычно этих ранних полуобезьян делят на лемуриформных (лемуры и их предки) и тарзииформных (долгопяты и их предки). Не позднее 40 млн лет назад, т. е., скорее всего, в середине или даже начале эоцена, происходит, судя по находкам в Восточной Азии и Северной Африке, обособление линии высших приматов — антропоидов (Anthropoidea), или, иначе говоря, собственно обезьян (рис. 4).
Рис. 4. Эволюционная ветвь высших приматов (Anthropoidea). Показаны лишь линии, представители которых сохранились поныне
Следует иметь в виду, что в отечественной литературе термин антропоиды часто применялся, а иногда применяется и сейчас, для обозначения человекообразных обезьян. Однако такое его употребление, хотя и находит некоторое оправдание в этимологии самого слова «антропоиды» (оно происходит от греческого άνθρωπος — человек и в буквальном переводе означает «человекоподобные»), нежелательно и ведёт к путанице. Антропоиды, согласно общепринятой зоологической номенклатуре, — это все обезьяны вообще, а для отдельного обозначения человекообразных обезьян существует и используется во всём мире термин «гоминоиды» (от латинского homo — человек).
Вопрос о том, где появились первые обезьяны, пока не решён. На роль их прародины претендуют Африка и Восточная Азия, но сделать выбор между этими регионами при нынешнем состоянии наших знаний не представляется возможным. В последние годы некоторые важные находки, способные пролить свет на проблему происхождения антропоидов, были сделаны на территории Китая, Бирмы и сопредельных с ними стран, хотя и Африка, наверняка, не сказала ещё своё последнее слово в споре континентов, и здесь палеонтологов тоже ждут новые открытия.
Уже первые обезьяны заметно отличались от низших приматов, или, иначе говоря, полуобезьян многими чертами строения зубной системы и глазничной области черепа, свидетельствующими об изменении характера рациона в сторону большей растительноядности (точнее, фругиворности, т. е. питания фруктами) и переходе от ночного образа жизни к дневному. С этими особенностями в их поведении связано и то обстоятельство, что зрение у них развито гораздо лучше, чем у большинства животных, а обоняние, наоборот, утратило остроту. По последним двум признакам к обезьянам близки долгопяты, на основании чего обе группы объединяют иногда в подотряд гаплорин (απλός по-гречески означает «простой», а ρίνος[2] — «нос»). Более важно, однако, что антропоидам присуща такая черта, как относительно большой размер мозга, чем долгопяты похвастаться не могут. У долгопятов, как и у остальных приматов, объём мозга примерно такой же, как у других животных аналогичного размера, тогда как у обезьян он в среднем в два раза больше, чем следовало бы ожидать от млекопитающих их «весовой категории».
Большинство палеонтологов выводят антропоидов из тарзииформных приматов, меньшинство — из лемуриформных, а, кроме того, высказывается предположение, что они могут представлять собой независимую ветвь, берущую начало непосредственно от общего для всех приматов исходного ствола. Впоследствии эта ветвь разделяется на две: широконосых обезьян (Platyrrhini), живущих сейчас только в Южной Америке, и узконосых (Catarrhini), населяющих Старый Свет. Наиболее древние кости широконосых обнаружены в Боливии, в отложениях возрастом около 25 млн лет, а вторых — в Северной Африке и на Аравийском полуострове, где они залегали в геологических слоях, сформировавшихся 31–35 млн лет назад, а частично, возможно, и ранее. Предки широконосых, бывшие, по-видимому, выходцами из Африки, могли случайно попасть в Южную Америку на естественных «плотах» из оказавшихся в воде растений. Сколь бы малой ни казалась вероятность благополучного исхода подобного путешествия, случаи такого рода, видимо, всё же происходили. Помимо предков широконосых обезьян, аналогичный путь проделали, как считается, и некоторые африканские грызуны.
Почти всё, что мы знаем сегодня о ранних стадиях эволюции узконосых обезьян, стало известно благодаря находкам в Файюмской впадине, расположенной к западу от Нила на территории Египта. Это крупнейшее местонахождение среднеолигоценовой фауны дало останки множества видов приматов. Они происходят из отложений геологической формации Джебел Катрани, относящихся к периоду от 31 до 35 млн лет назад. Наибольший интерес среди обезьян, кости которых были найдены в Файюме, имеют для нас так называемые проплиопитециды, выделяемые обычно как надсемейство. К числу проплиопитецид относят роды проплиопитека (Propliopithecus) и египтопитека (Aegyptopithecus). В них многие исследователи видят наиболее вероятных предков современных узконосых обезьян, в том числе и человекообразных.
Происхождение и эволюция человекообразных обезьян
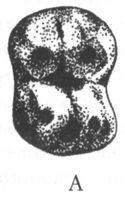
Примерно на рубеже олигоцена и миоцена (23 млн лет назад), или чуть раньше (см. рис. 2) происходит разделение дотоле единого ствола узконосых обезьян на две ветви: церкопитекоидов, или собакоподобных (Cercopithecoidea) и гоминоидов, т. е. человекообразных (Hominoidea). Это разделение, по всей видимости, во многом было обусловлено тем, что часть узконосых (предки церкопитекоидов) перешла к питанию листьями, тогда как другая часть (предки гоминоидов) сохранила верность фруктовой диете. Различия в меню сказались, в частности, на строении зубов, что чрезвычайно важно для палеонтологов, поскольку именно зубы составляют бо́льшую часть ископаемых находок. Поверхность жевательных зубов церкопитекоидов имеет характерный, только им присущий узор, образуемый четырьмя бугорками. На зубах же человекообразных обезьян пять округлых бугорков, разделённых У-образной бороздкой — так называемый «узор дриопитека» (рис. 5).
Рис. 5. Поверхность коренных зубов церкопитекоидов (А) и гоминоидов (Б)
Церкопитекоидов, представленных единственным, но очень многочисленным семейством мартышкообразных, часто называют низшими узконосыми обезьянами, а гоминоидов — высшими. Помимо особенностей формы зубов, гоминоидов отличает от низших узконосых обезьян также отсутствие хвоста, более короткое (по отношению к конечностям), плоское и широкое туловище и, наконец, специфическое строение плечевого сочленения, обеспечивающее большую свободу вращения верхних конечностей в разных плоскостях. По-видимому, все перечисленные признаки были приобретены ранними гоминоидами в результате приспособления к таким способам передвижения по деревьям, которые требуют вертикального и хотя бы частично выпрямленного положения тела. Это лазание с опорой на нижние конечности, а также так называемая брахиация, т. е. перенос или перебрасывание тела с ветки на ветку с помощью верхних конечностей (рис. 6). Для низших обезьян ни то, ни другое, в общем, не свойственно, и они, в отличие от человекообразных, даже по ветвям передвигаются, как правило, на четырёх конечностях, подобно всем остальным млекопитающим от белки до леопарда.
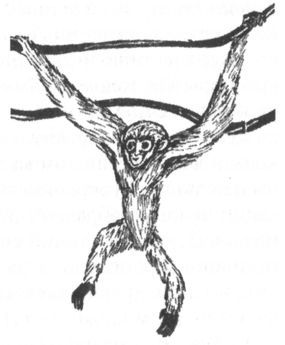
Рис. 6. Гиббоны — классические брахиаторы
Одно время некоторые исследователи полагали, что церкопитекоиды и гоминоиды разделились ещё в раннем олигоцене, и что уже проплиопитеки и египтопитеки, жившие примерно 30–35 млн лет назад, должны рассматриваться как гоминоиды. Действительно, зубы этих обезьян, найденные в Файюмской впадине, несут хорошо выраженный узор дриопитека, но вот кости их черепа и скелета по своему строению ближе к аналогичным костям церкопитекоидов. Такая мозаичность признаков позволяет видеть в этих родах более или менее близкое подобие той предковой формы, от которой произошли церкопитекоиды и гоминоиды. К сожалению, огромный временной интервал, захватывающий весь поздний олигоцен, всё ещё остаётся практически неохарактеризованным ископаемым материалом, и поэтому представить себе сколько-нибудь детально процесс расхождения двух ветвей узконосых обезьян пока невозможно.