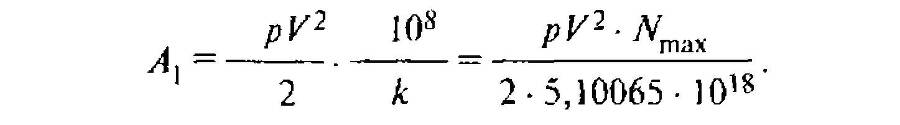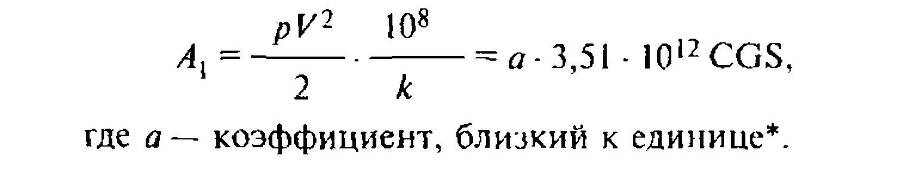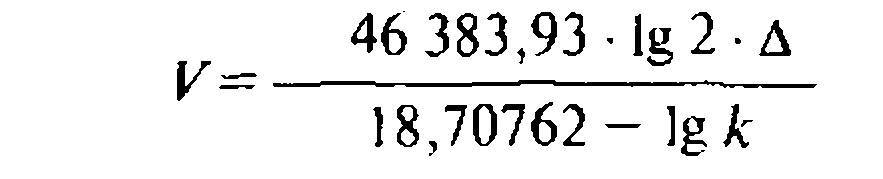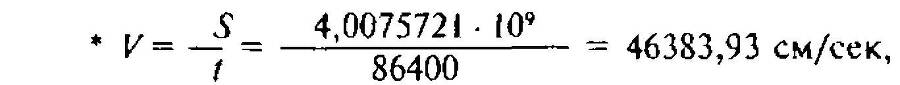§39. Скорость передачи жизни может давать нам ясное понятие о геохимической энергии жизни разных организмов. Она колеблется в больших пределах и находится в тесной зависимости от размера организма. Для самых мелких организмов, для бактерий, она, как мы видели, близка к скорости звука, т. е. к величине, равной 33 100 см/с. Для самых крупных, для крупных млекопитающих, она равна долям сантиметра — для индийского слона, например, К— 0,09 см/с.
Это крайние пределы. Между ними помешаются скорости передачи жизни для всех других организмов. Они находятся в явной зависимости от размеров организма и в более простых случаях (например, для организмов, форма которых приближается к шару) связь размеров организма с его скоростью v может быть уже сейчас математически выражена.
5,10064 • 108
-= 13 963,3, где 5,10064 • 10х км2 — поверхность Земли. — Прим. ред.
365
Но существование определен ной математической зависимости всегда и везде в этой области несомненно и отвечает старинному прочному эмпирическому обобщению (§32).
§ 40. Скорость передачи жизни дает ясное понятие об энергии жизни в биосфере, о ее в ней работе, но оно недостаточно для ее определения. Для этого мы должны принять во внимание массу того организма, энергия растекания совокупностей которого в биосфере определяется скоростью V
Выражение pVу/2 , где р— средний вес организма1,
скорость растекания геохимической энергии которого равна V, даст нам выражение кинетической геохимической энергии живого вещества. Взятое по отношению к определенным площади или объему биосферы, оно может дать нам выражение той химической работы, которая в геохимических процессах этой плошади или объема может быть произведена данным видом или расой организмов.
Уже давно мы имеем подходы к определению этим путем по отношению к определенной плошади биосферы — гектару — части геохимической энергии живого вещества. Это делается при определении урожаев — количества с данной площади полезных человеку организмов или их продуктов.
В более полной форме оно выражается в количестве органического вещества, которое может быть создано размножением и ростом организмов на гектаре.
Хотя эти данные очень неполны и не охвачены теорией в достаточной степени, они привели уже к важным эмпирическим обобщениям.
Несомненно, количество создаваемого на гектаре органического вещества ограничено и теснейшим образом связано
с той солнечной лучистой энергией, которую захватывает зеленое растение. Геохимическая энергия, собранная этим путем — размножением организмов, — есть измененная солнечная энергия.
Во-вторых, все более выясняется, что в случаях максимальных урожаев количества органического вещества с гектара почвы и гектара океана суть числа одного порядка, приближающиеся к одной и той же величине. Гектар суши охватывает ничтожный слой, не превышающий метров, гектар океана отвечает слою воды, захваченной жизнью, измеряемому километрами. Тождественность создаваемой в них энергии жизни, очевидно, указывает на освещение сверху как на ее источник.
Мы увидим, что, вероятно, это связано с характерным свойством почвы суши, скопляющей в себе концентрацию организмов, обладающих огромной геохимической энергией (§ ! 55). Благодаря этой концентрации энергии живого вещества тонкий слой почвы по своему геохимическому эффекту может сравниваться с огромной толщей моря, где центры жизни разжижены инертной массой воды.
§41. Кинетическая геохимическая энергия организма
pV2/2, взятая по отношению к гектару, т. е. к 108 см2, выражается следующей формулой, где108/k — количество организмов на гектар при достижении ими стационарного числа (§ 37), а к — коэффициент плотности жизни (§ 36):
Чрезвычайно характерно, что эта величина для Protozoa есть величина постоянная. Для них всех выражение A1 принимает форму:
Отвечает плохо известному удельному весу Protozoa.
Из этой формулы ясно, что кинетическая геохимическая энергия Protozoa определяется скоростью К связанной с весом, размерами организма и темпом размножения Д. Отнесенное к ∆, V выражается следующей простой формулой*:
в которой числовые, постоянные для всех видов организмов коэффициенты связаны с размерами планеты (с плошадью ее поверхности и длиной ее экватора, причем величины выражены в сантиметрах и секундах)**.
Из формулы скорости очевидно, что одни размеры планеты не могут объяснить действительно существующего предела для V и ∆.
Наибольшая нам известная величина V равна 33 100 см/с, а наибольшая величина ∆ — около 63—64.
Могут ли они идти дальше (что, как это видно из приведенных формул, возможно и при постоянстве кинетической энергии на гектар) или есть в биосфере условие, этому препятствующее?
Такое условие есть, и им является газовый обмен организмов, неизбежный и необходимый для их существования и, в частности, для их размножения.
§ 42. Организм не существует без газового обмена, без дыхания. Чем размножение идет быстрее, тем дыхание становится интенсивнее. По степени газового обмена мы всегда можем судить об интенсивности жизни.
В масштабе биосферы нам необходимо, конечно, иметь в виду не дыхание отдельного организма, а общий результат дыхания, необходимо учесть газовый обмен — дыхание всех живых организмов, обнять его как часть механизма биосферы.
длина экватора, t — число секунд в сутках; 18 lg 5,10065 =18,70762. — Прим. ред.
** Выражение V существует для всех организмов, а не только для Protozoa. Формула А1 для высших групп, для Methazoa и Metaphyta имеет иную, меньшую величину, что связано с явлениями дыхания и глубоким отличием их организации от организации простейших. Не могу здесь входить в рассмотрение этих важных и сложных явлений.
В этом отношении давно имеются эмпирические обобщения, очень мало до сих пор обращавшие на себя внимание и не учтенные на щей научной мыслью.
Одно из них указывает, что газы биосферы те же, которые создаются при газовом обмене живых организмов. В биосфере существуют только они одни: О2 N2, СO2, Н2О, Н2, СН4, NH3... Это не может быть случайностью.
Затем весь свободный кислород биосферы создается на земной поверхности только благодаря газовому обмену зеленых организмов. Этот свободный кислород есть главный источник свободной химической энергии биосферы.
И, наконец, в-третьих, количество этого свободного кислорода в биосфере, равное 1,5 • 1021 г, есть число того же порядка, как и количество существующего и с ним неразрывно связанного живого вещества, исчисляемого в 1020—1021 г. Оба эти исчисления получены вполне независимо друг от друга.
Эта тесная связь газов Земли с жизнью указывает с несомненностью, что газовый обмен организмов — и на первом месте их дыхание — должен иметь первостепенное значение в газовом режиме биосферы, т. е. являться планетным явлением.
§ 43. Этот газовый обмен — дыхание — определяет весь темп размножения; он ставит пределы для величин V и ∆.
Они не могут перейти через пределы, нарушающие свойства газов.
Я уже указывал (§ 29), что количество организмов, могущих существовать в одном кубическом сантиметре среды, должно быть меньше количества в нем газовых молекул, т. е. меньше 2,706- 1019 (число Лошмидта). Если величина v будет больше 33 100 см/с, то для организмов, размеры которых меньше размеров бактерий (т. е. размеров меньшего порядка, чем п • 10-5), количество неделимых благодаря их размножению превысит 1019 в 1 см3. Очевидно, при неизбежном существовании обмена между газовыми молекулами и организмами количество поглощающих и выделяющих газовые молекулы организмов, соизмеримых с молекулами по своим размерам, должно было бы расти по мере уменьшения размеров организмов с все большей, в конце концов, невероятной быстротой.