Поэтому хвост за спиной, который не используется в качестве руля, был бы помехой для гиббона, который передвигается с помощью вертикальной брахиации (а также, предположительно, и для сопредка № 4).
Вот и все, что я могу об этом сказать. Думаю, зоологам нужно уделить больше внимания выяснению того, почему мы, человекообразные обезьяны, утратили хвост. Ведь возникает масса проблем! Например, как хвост сочетался бы с обычаем носить одежду, особенно брюки? Это придало бы новый смысл традиционному вопросу портного: “Сэр, вам ширинку налево или направо?”
Рассказ Гиббона[12]
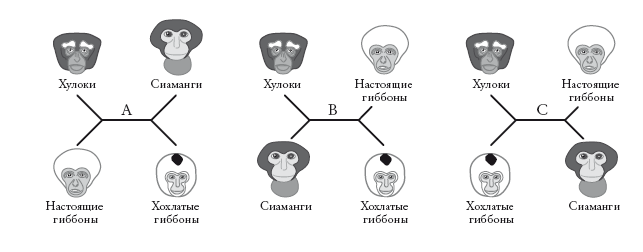
На рандеву № 4 мы встречаем крупную группу пилигримов. И теперь могут возникнуть проблемы с установлением родства. (Чем дальше, тем затруднительнее это сделать.) Существует двенадцать видов гиббонов, принадлежащих к четырем основным группам. Это Bunopithecus (группа, представленная одним видом, известным как хулок); настоящие гиббоны Hylobates – шесть видов, самый известный – белорукий гиббон (Hylobates lar); сиаманг (Symphalangus) и номаски (Nomascus) – четыре вида “хохлатых” гиббонов. Сейчас я объясню, как построить схему эволюционных отношений, или филогению, для этих четырех групп.
Генеалогические деревья могут быть укорененными или неукорененными. В случае укорененного древа нам известно, кто является предком. Большинство деревьев в этой книге – укорененные. Неукорененные деревья, напротив, не отражают направление эволюции. Их называют звездчатыми диаграммами. В них не заложена стрела времени, и нельзя сказать, где у них начало, а где конец. Здесь приведены три примера, описывающие отношения четырех родственных групп гиббонов.
Неважно, какая ветвь окажется справа в точке ветвления, а какая – слева. Длина ветвей до сих пор не имела значения (это скоро изменится). Древовидная диаграмма, в которой длина ветвей не несет информации, называется кладограммой (в данном случае неукорененной). Порядок ветвления – вот главная информация, отраженная в кладограмме. Попробуйте перевернуть любую из боковых вилок вокруг центральной горизонтальной линии: это ничего не изменит в схеме отношений между группами.
Эти три неукорененные кладограммы описывают возможные отношения четырех видов – при условии, что мы рассматриваем лишь случаи дихотомического ветвления. Как и для укорененных деревьев, случаи разделения на три (трихотомия) и больше ветвей (политомия) мы допускаем, когда у нас недостаточно информации (“неразрешенные” ветви).
Любая неукорененная кладограмма может стать укорененной – для этого нужно указать самую старшую точку на древе (“корень”). Некоторые исследователи – те, на которых мы ссылались при рассмотрении древа в начале этого рассказа, – предлагают для гиббонов укорененную кладограмму слева. Другие предпочитают укорененную кладограмму справа. На первой схеме хохлатые гиббоны (номаски) представлены дальними родственниками всех остальных гиббонов. На второй схеме на их место помещен хулок. Несмотря на это различие, оба древа производны от одного неукорененного дерева (А). Кладограммы отличаются лишь корнем. На первой он расположен на ветви номасков, а на второй – на ветви Bunopithecus.
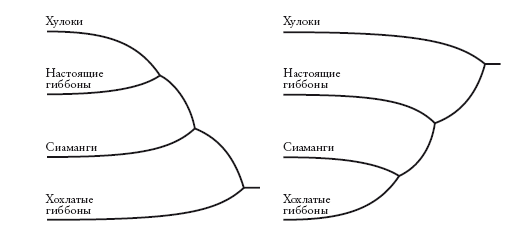
Как происходит “укоренение”? Самый распространенный способ – расширить древо, включив в него по меньшей мере одну “внешнюю” группу, которая является заведомо далеким родственником по отношению ко всем другим представленным группам. Например, на древе, построенном для гиббонов, внешней группой может быть орангутан или горилла – а еще лучше слон или кенгуру. Можно сколько угодно сомневаться по поводу взаимоотношений групп гиббонов, но мы точно знаем, что общий предок любого гиббона с большими человекообразными обезьянами (или слоном) старше, чем общий предок любого гиббона с любым другим гиббоном. Поэтому, строя древо, включающее гиббонов и крупных человекообразных обезьян, мы не ошибемся, поместив корень где-то между ними.
Легко заметить, что три неукорененных древа, которые я нарисовал, описывают все возможные дихотомические деревья для четырех групп. Для пяти групп будет 15 таких деревьев. Но не стоит и пытаться сосчитать количество возможных деревьев для, скажем, 20 групп. Их сотни миллионов миллионов миллионов[13]. Число резко возрастает с ростом числа групп, которые мы желаем классифицировать, и даже у самого мощного компьютера такие расчеты могут занять целую вечность. Однако в принципе задача довольно проста. Из всех возможных деревьев нужно выбрать те, которые лучше всего объясняют сходства и различия наших групп.
Но что значит – “лучше всего объясняют”? Когда мы рассматриваем выборку животных, количество сходных и отличных черт может оказаться практически бесконечным. Сосчитать их труднее, чем кажется. Нередко один “признак” является неотделимой частью другого. И если мы сочтем эти признаки независимыми, окажется, что на самом деле мы учли одни и те же признаки дважды. Представьте, например, многоножек четырех видов: A, B, C и D. Многоножки A и B сходны во всем, кроме того, что у А конечности красные, а у B – синие. Многоножки C и D сходны друг с другом и отличаются от A и B – но у C конечности красные, а у D – синие. Если мы сочтем цвет конечностей одним “признаком”, мы справедливо поместим A и B в одну группу, а C и D – в другую. Но если мы будем считать каждую ножку из ста отдельным признаком, количество этих признаков перевесит все остальные, и тогда A сгруппируется с C, а B – с D. Очевидно, что в этом случае мы просто сто раз посчитали один и тот же признак. А на самом деле это один признак, потому что цвет всех ста ножек определяется одним эмбриологическим “событием”.
То же верно и для двусторонней симметрии: эмбриогенез таков, что, за редкими исключениями, одна сторона тела животного является зеркальным отражением второй. Ни один зоолог, строя кладограмму, не будет считать дважды “левый” и “правый” признак. Впрочем, не всегда очевидно, какие признаки независимы. Голубю нужна крупная грудина для крепления летательных мышц. А нелетающим птицам, например киви, она не нужна. Должны ли мы считать мощную грудину и способные к полету крылья двумя независимыми признаками, отличающими голубя от киви? Или сочтем их единым признаком на том основании, что состояние одного признака определяет состояние второго – или, по крайней мере, уменьшает его изменчивость? В случае многоножек и зеркальной симметрии правильный ответ очевиден. А в случае грудины – нет. На этот счет может иметься две вполне обоснованные точки зрения.
До сих пор мы говорили о внешнем сходстве и различии. Однако внешние признаки эволюционируют лишь в том случае, если они – проявления последовательностей ДНК. Сегодня мы можем непосредственно сравнить последовательности ДНК. Дополнительное преимущество ДНК заключается в том, что она имеет длинные цепочки, и “текст” ДНК предоставляет гораздо больше признаков, которые можно считать и сравнивать. Проблемы крыльев и грудин просто тонут в огромном потоке данных, которые дает нам ДНК. Более того, многие различия в ДНК “невидимы” для естественного отбора и поэтому являются более “чистыми” свидетельствами родства. Например, многие сочетания нуклеотидов в ДНК синонимичны: они кодируют одну и ту же аминокислоту. Мутация, меняющая сочетание нуклеотидов на синонимичное, невидима для естественного отбора. Однако для генетика такая мутация не хуже любой другой. То же относится и к “псевдогенам” (обычно это случайные копии работающих генов), и ко многим другим “мусорным” последовательностям ДНК, которые располагаются на хромосомах, но не считаются и не используются. Независимая от естественного отбора ДНК получает возможность свободно мутировать, а это обеспечивает специалистов по систематике высокоинформативными данными. Это не отменяет того, что некоторые мутации могут иметь реальный и значительный эффект. Их замечает отбор, они отвечают за видимую глазу красоту и сложность всего живого.