Можно было продолжить рассмотрение материалов по географическому распространению проблематичных фосфатных организмов, называемых в специальной литературе SSF (Small shally fossils — мелкие скелетные ископаемые), но, вероятно, это не имеет уже больше смысла. Все они достаточно относительны. Нам осталось лишь упомянуть всего одну группу — хиолительминты. Их принадлежность к седентарным (прикрепленным ко дну) организмам как будто не вызывает ни у кого никаких сомнений. Но именно эта группа встречена повсеместно там, где мы сталкиваемся с нижнекембрийскими, особенно томмотскими, отложениями.
В заключение краткого обзора биогеографических данных следует сказать несколько слов о биогеографии по водорослям. Я уже упоминал, что карбонатные водоросли географически распространены были там же, где и археоциаты, но в ряде случаев и еще шире, практически во всех районах, где мы встречаем достоверные нижнекембрийские известняковые отложения. Состав водорослей почти не отличим по всему миру, и точно так же, как и для археоциат, их наибольшее разнообразие известно в настоящее время из складчатого обрамления Сибирской платформы. Наиболее же популярны роды Renalcis, Girvanella, Epiphyton.
Теперь попробуем рассмотреть некоторые вопросы, связанные с изучением центров происхождения кембрийских фаун и флор.
Различными методами было установлено, что большинство групп, известных в раннем кембрии, приобретает возможность строить скелет в геологическом смысле одновременно. Подчеркнем еще раз, что это именно в геологическом смысле, потому что точность наших построений ±1 млн. лет. Но и такая точность огромна.
В результате момент появления возможности строить скелет у большого числа разнообразных групп является прекрасным хронологическим репером. Теперь вспомним, что целый ряд групп именно только с этого времени фиксируется нами в скелетном виде. И, таким образом, с небольшими допущениями мы можем считать это временем первого появления данной группы в том виде, в каком мы встречаем ее затем в фанерозое. Это дает нам огромные возможности для распознания центров происхождения многих групп.
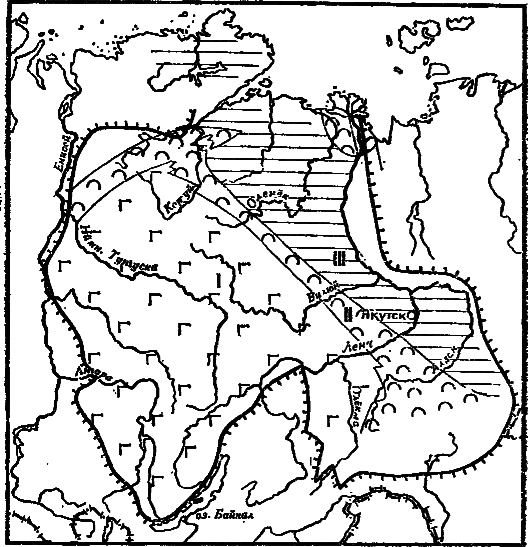
Рис. 41. Схема положения «переходной зоны» (II), разделяющем «лагунный» (I) и открыто морской (III) бассейны Сибирской платформы «Переходная зона» — центр происхождения ряда групп раннекембрийских фаун
Наиболее изученными и поэтому наиболее показательными в этом отношении являются, конечно, археоциаты. Самые древние из известных в мире археоциат обнаружены на Сибирской платформе в самых низах томмотского яруса. Более того, они обнаружены в пределах очень узкой полосы, пересекающей Сибирскую платформу с юго-востока на северо-запад. Ширина этой полосы обычно несколько десятков километров, а протяженность — более 2000 км (рис. 41). Она разделяет два крупных совершенно разных по характеру бассейна. На запад от нее располагался обширный бассейн с явно нарушенной (повышенной) соленостью вод, где часто образовывались доломиты, гипсы и даже соли. К востоку же располагался открыто морской бассейн с нормально солеными водами и, вероятно, несколько большими глубинами. Этот восточный бассейн, вероятнее всего, открывался в «океан». Уже в пределах этой полосы, или так называемой «переходной зоны», можно локализовать наиболее древний район с археоциатами, который расположен на юго-востоке Якутии в бассейне среднего течения р. Алдан. Вероятно, именно этот район и является центром происхождения этой группы.
Первые археоциаты начали совместно с водорослями строить небольшие тела размером не более 1 м в диаметре, называемые биогермами. До этого момента способность к биогермостроению была только у бактерий и водорослей (строматолитов). Если наши выводы верны, то несомненно интересно, в каких же условиях появились первые археоциаты.
Очевидно, это не только крайнее мелководье (первые метры), но и достаточно высокие температуры воды. Мы уже говорили о том, что, по оценкам специалистов, температуры воды должны были быть около 25—30°, т. е. это те условия, при которых сейчас формируются коралловые рифы. Очень важным элементом обстановки, в которой формировались первые археоциаты и археоциатоводорослевые биогермы, является характер осадконакопления. А осадки накапливались здесь карбонатные, причем исключительно Ca-карбонатные, т. е., в переводе на обычный язык, на дне накапливались известковые илы, которые позднее превращались в известняки.
Как ни странно, характерной особенностью этих осадков является и повышение содержания железа, в результате чего почти все древнейшие томмотские известняки Сибирской платформы, где обнаружены археоциаты, имеют красную окраску.
Дальнейшая судьба археоциат, естественно, должна была зависеть от степени распространенности подобных обстановок на земном шаре. Несколько огрубляя проблему, можно сказать, что их судьба зависела от того, сколь во многих местах известняки будут накапливаться на планете. В течение томмотского времени археоциаты сначала осваивали «переходную зону», достигнув ее северного окончания через 0,5—1,0 млн лет, т. е. средняя скорость миграции составляет 2—4 км в 1000 лет).
В конце томмотского века и начале атдабанского археоциаты сделали попытку вырваться за пределы Сибирской платформы, И это увенчалось успехом лишь в связи с развитием вокруг более или менее подходящих обстановок, в которых археоциаты смогли прижиться.
Надо сказать, что при освоении новых районов и несколько иных экологических ниш (условий) возникали новые центры видообразования, которые не совпадали с первичным центром происхождения археоциат. Так, в атдабанское время возник крупнейший центр видообразования археоциат — Алтае-Саянская складчатая область, которая просуществовала как центр видообразования до самого конца раннего кембрия.
В позднеатдабанское время и в начале ботомского формируются еще несколько центров видообразования. Это прежде всего Северная Африка (Марокко), запад Северной Америки (уже упоминавшаяся Американо-Колымская область) и Южная Австралия. Исследования по археоциатам подтвердили выводы акад. Н. И. Вавилова, сделанные на примере злаковых растений более полувека назад, о том, что центры происхождения и диверсификации чаще всего могут не совпадать.
Надо сказать, что археоциаты достаточно детально исследовались с этой точки зрения (с точки зрения центров их происхождения). Что же касается других групп, эта задача практически никогда не ставилась. Однако по многим из групп уже сейчас могут быть сделаны однозначные выводы или поставлены очень конкретные вопросы, решение которых прольет свет на проблему центров происхождения групп. Приведу несколько примеров.
Сначала, естественно, обсудим состояние дел с рядом групп, появившихся совместно с археоциатами в начале томмотского века в пределах «переходной зоны» Сибирской платформы. Отсюда известны: губки (разрозненные спикулы), моллюски, брахиоподы и различные фосфатные проблематики (SSF) и т. д.
Относительно губок вопрос очень сложен. Существуют многочисленные указания на наличие спикул губок в докембрии. Подавляющее большинство таких указаний ошибочно и связано с неверной диагностикой спикул губок или неточным определением возраста пород. Однако часть указаний на сегодняшний день по разным причинам непроверяема. В связи с этим абсолютно исключить возможность появления спикул у губок в докембрийское время невозможно, и соответственно сегодня вопрос о центре пли центрах происхождения губок решен быть не может даже в самом приблизительном варианте.
Значительно лучше дело обстоит с древнейшими моллюсками. Более или менее очевидно, что «переходная зона» и прилегающие к ней зоны открытого моря несомненно были районом происхождения алданелл (Aldanella) и ряда других форм гастропод (или частично, возможно, моноплакофор). Расселение их происходило довольно быстро, во всяком случае алданелл (не исключен их планктонный образ жизни). В начале зоны D. regularis (второй зоны томмотского яруса) алданеллы достигли Ньюфаундленда. Исходя из сегодняшнего положения Сибирской платформы и Ньюфаундленда, средняя скорость миграции должна быть соизмерима со скоростью миграции археоциат — около 5 км в 1000 лет (а если исходить из наших реконструкций положения континентов, то и того меньше — около 3,5—4,0 км в 1000 лет).