Вот пример, малоизвестный и убедительный. Наш великий садовод И. В. Мичурин в 1915 году писал: «В последнее время наши неофиты дела гибридизации как-то особенно назойливо стараются нам навязать этот гороховый закон — создание австрийского монаха». Однако Мичурин был настоящим ученым и решил проверить законы Менделя на гибридах обычной яблони с яблоней Недзвецкого. Последний вид — очень удачный партнер для скрещивания: у него красные плоды, красная кора и даже хлорофилл листьев замаскирован красным антоциановым пигментом. Уже в 1917 году Мичурин описал первые подобные опыты, где были получены четкие числовые расщепления, а двадцать лет спустя отмечал: «В законе Менделя я нисколько не отвергаю его достоинств» — и рекомендовал для показательных скрещиваний, демонстрирующих применимость менделизма в генетике плодовых, именно яблоню Недзвецкого, а также краснолистную грушу, сливу и орех. Поэтому было бы кощунством утверждать, что Мичурин до конца дней отвергал менделизм. Все обстояло как раз наоборот.
Колоссально много для развития учения о наследственности сделал американец Томас Гент Морган, который вначале был столь же ярым противником Менделя и решил его опровергнуть на другом объекте — кроликах. Однако попечители Колумбийского университета, где работал Морган, сочли кроликов чересчур дорогими; Морган не пал духом и использовал для опытов крошечную плодовую мушку — дрозофилу. Дрозофила стала классическим объектом генетики — редчайший случай, когда скупость снабженцев оказала науке неоценимую услугу! Дело в том, что за год можно получить 25 поколений дрозофил и разместить на одном лабораторном столе их многотысячные «стада». Помимо этого, в работе с дрозофилой есть какая-то непонятная, но властная притягательность. Я помню, как мне в свое время пришлось поставить на ней несколько опытов — я без особой нужды для основной работы затянул их на добрые полгода и прервал с великим сожалением. Это, как говорят на лабораторном жаргоне, болтливый объект — так много из него познаешь нового.
Рассказать о всех открытиях Моргана и созданной им научной школы здесь невозможно: для этого потребовалась бы книга по меньшей мере такого же объема. Да в этом и нет нужды — в последнее время вышло достаточно хороших популярных книг, посвященных успехам генетики. Мы рассмотрим лишь те открытия, которые имеют первостепенное значение для эволюционной теории.
Морган и его не менее прославившиеся ученики — А. Стертевант, Г. Мёллер и К. Бриджес в 1909–1911 годах доказали, что второе правило Менделя не абсолютно. Полной независимости в распределении наследственных задатков нет: они распределены по группам сцепления, иными словами, — могут передаваться потомству лишь совместно. У дрозофилы четыре таких группы — столько же, сколько пар хромосом в клетках тела, и столько же, сколько хромосом получает мужская или женская половая клетка — гамета. Гениальное предвидение Августа Вейсмана, в XIX веке связавшего наследственность с хромосомами, полностью подтвердилось.
Далее, само правило сцепления оказалось неабсолютным. Гены могли переноситься из одной хромосомы в другую. Для тех, кто забыл школьную биологию или учился в период с 1948 по 1964 годы, напомним, что при делении клетки, приводящем к образованию гаметы (мейоз), хромосомы не делятся. Парные, гомологичные хромосомы выстраиваются друг против друга, а затем расходятся по клеткам. В результате яйцеклетка и спермий имеют уменьшенное вдвое число хромосом — у человека — 23, у дрозофилы — 4 (так называемый гаплоидный набор). Лишь слияние гамет, приводящее к возникновению оплодотворенной яйцеклетки — зиготы, восстанавливает прежний, диплоидный набор, характерный для вида.
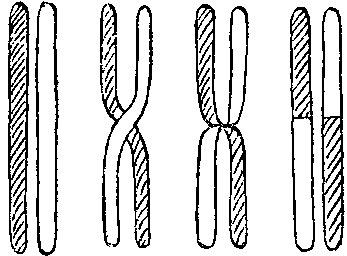
Группа Моргана показала, что, выстроившись попарно, гомологичные хромосомы при мейозе могут обменяться частями. Это явление было названо кроссинговером (перекрестом). Оказалось, что цитологи уже давно наблюдали кроссинговер в клетках многих организмов.
Анализ кроссинговера показал, что гены в хромосомах расположены линейно в одномерной последовательности, как буквы в строке. Это позволило построить хромосомные карты генных локусов (локус — по-латыни просто место, в генетике — место, где расположен в хромосоме ген. Каждый локус может быть занят аллелем — формой какого-либо гена — например, аллелем желтой или зеленой окраски семядолей гороха). Чем дальше в хромосоме отстоят друг от друга локусы, тем больше вероятность того, что они расстанутся при кроссинговере.
Для эволюции кроссинговер имеет первостепенное значение. Возьмем для примера менделевский горох. Допустим, что ген желтой окраски горошин и ген, обуславливающий развитие длинных корней, находятся в одной хромосоме. Не будь кроссинговера, получение гороха с зелеными горошинами и длинными корнями было бы невозможным. Кроссинговер делает сцепление между генами неабсолютным: в результате его оба признака могут оказаться в одной хромосоме. Еще более важен для возникновения новых форм эффект неравного кроссинговера, открытый в 1925 году учеником Моргана Стертевантом. Стертевант изучал ген, вызывающий появление у дрозофилы полосковидных глаз, и установил, что возможны случаи не абсолютно точного обмена частями хромосом. После такого неравного кроссинговера в одной хромосоме оказывается два одинаковых локуса, а в другой — ни одного. Один и тот же локус, представленный двумя аллелями, становится двумя самостоятельными одноаллельными локусами. У многих организмов одинаковые гены многократно дублированы: в хромосоме имеется не один локус, а целая цепочка их. В таких случаях неравный кроссинговер — явление весьма частое: одна хромосома получает львиную долю локусов, другая — меньше их число. Идет как бы переселение генов из одной гомологичной хромосомы в другую. Однако возможен и обратный процесс выравнивания числа локусов: кроссинговер в этом случае восстанавливает равенство.
Первые успехи генетики позволили понять механизм комбинаторной, или комбинативной, изменчивости, имеющей огромное значение в эволюции. При мейозе отцовские и материнские хромосомы, образующие пары, расходятся по гаметам случайно. Каково же число возможных сочетаний пар хромосом? Как показывает математика, оно равняется 2 в степени, равной числу пар гомологичных хромосом. У кролика, например, оно равно 222 = 4 385 000, у дрозофилы 24 = 16. Остается еще раз восхвалить скупость попечителей Колумбийского университета. Используй Морган кроликов вместо дрозофилы, он мог бы просто утонуть в море генетических рекомбинаций, и группы сцепления были бы открыты позже.
Если мы учтем перетасовывающую деятельность кроссинговера, нам станет ясно: запасы комбинаторной изменчивости практически неисчерпаемы. Поэтому в природе не может быть двух размножающихся половым путем организмов с идентичными наборами генов, нет двух одинаковых особей. Исключение (подтверждающее правило!) — однояйцовые близнецы, черенки с одного куста, бактерии, полученные в потомстве от одной клетки.
Представим некий организм, у которого в хромосомах всего 1000 локусов (на деле даже у бактерий эта величина больше). Если каждый локус представлен серией из 10 аллелей, число возможных сочетании из них — 101000 (как указал английский генетик С. Райт, эта величина неизмеримо больше числа электронов во всей Вселенной). Разумеется, львиная доля подобных сочетаний была бы нежизнеспособной при любых изменениях среды. Но и того, что остается, более чем достаточно, чтобы служить «сырьем» для эволюционного процесса, поставляя отбору неисчерпаемый материал в течение практически бесконечного времени.
Гены влияют друг на друга
Генетические карты, построенные морганистами, выглядели на первый взгляд странно. Гены, определяющие развитие самых различных признаков, мирно уживались в хромосоме рядом, как бусинки на нитке. Могло сложиться впечатление (а у некоторых оно и сложилось), будто организм — не что иное, как мозаика признаков, каждый из которых независим от других и определяется одним геном.