Рассмотрим данное положение на нескольких примерах. Один из них приводит Кольцов — это параллельная эволюция хищников и жертв — мезозойских динозавров. На протяжении многих миллионов лет триасового, юрского и мелового периодов среди травоядных ящеров шел жесткий отбор на увеличение размеров, ускорение роста. Более быстрорастущие и достигающие больших размеров ящеры быстрее выходили из-под «пресса хищников» (само слово «пресс» здесь не совсем удачно, действие хищников на популяции можно сравнить скорее с насосом). Однако и среди хищников шел параллельный отбор на возрастание размеров и мощи. Травоядные динозавры «ответили» освоением новых зон обитания — широкой адаптивной радиацией, переходом к земноводному образу жизни, а оставшиеся на суше развили сложные и мощные оборонные структуры. К концу мелового периода эти тенденции достигли предела. Появились настоящие «живые танки» — анкилозавры — огромные и неповоротливые, требующие массу пищи и покрытые сплошной костяной броней. У других динозавров — цератопсид возникли мощные многометровые рога. Хищники также достигли своего предела — появились гигантские тиранозавры и карнозавры, ходившие на двух ногах, напоминавших ноги чудовищной хищной птицы, череп их неимоверно увеличился. Огромные зубы-кинжалы могли закалывать жертву любых размеров. Многие из этих «конструктивных решений» носили характер явной спешки, несогласованности. Передние лапы тиранозавра превратились, например, в крошечные бесполезные для добычи пищи придатки, которыми нельзя было даже поковырять в зубах.
Эта система находилась в явно неустойчивом равновесии. Сравнительно небольшое похолодание, изменившее характер растительности, привело к гибели от голода травоядных ящеров. За ними последовали не нашедшие пищи хищники, и освободившуюся от динозавров Землю стали быстро заселять державшиеся до того в тени млекопитающие — мелкие ночные зверьки, которых гигантские ящеры даже не замечали.
А вот второй пример, так сказать, обратный по знаку. Принято считать, что мелкие размеры большинства грызунов — следствие перехода к подземному образу жизни в хорошо защищенных от хищников норах. Сопряженной, согласованной с изменениями своих жертв эволюции подвергся и самый мелкий хищник нашей фауны — ласка, которая может преследовать мышей в норах.
Иногда стратегия в игре «Вид против Среды» настолько прямолинейна, что эволюция выглядит неким направленным процессом — ортогенезом. Так пытались трактовать эволюцию современной лошади. Ее предок — эогиппус жил в условиях мягкого влажного климата эоценового периода, был четырехпалым, и его зубы были хорошо приспособлены к пережевыванию лесной и болотной растительности. Современная лошадь много крупнее эогиппуса, ее однопалые конечности приспособлены к быстрому продолжительному бегу, а зубы — к жесткой траве сухих степей. Построив ряд предков лошади во времени (нечто вроде родословной), мы наблюдаем направленные изменения конечностей, зубов и размеров тела. Однако, как оказалось, древние лошади развивались не по одному, а по нескольким направлениям, параллельными стволами и целыми букетами видов и форм. Все эти стратегии оказались неудачными, кроме одной, — той, которая привела к современной лошади И родословное древо ее меньше всего напоминает телеграфный столб, как полагают ортогенетики.
Какое значение имеет игра между популяцией и средой для прогресса? Самое прямое: чем сложнее среда, чем больше других видов организмов воздействуют на вид, тем более сложными становятся условия существования вида, поэтому он должен усложниться или вымереть.
Некоторую аналогию можно найти в эволюции оружия. Ясно, что самый лучший воин, типа д'Артаньяна, вооруженный шпагой, кремневым пистолетом и мушкетом, не выдержал бы поединка против любого новобранца современной армии с пистолет-пулеметом. Современный автомат намного сложнее мушкета, но отнюдь не потому, что конструкторы оружия любят сложность ради сложности.
Доказательство этого принципа можно найти от обратного, как говорят математики. В условиях упрощения среды потребности в прогрессивной эволюции уменьшаются. Именно поэтому мы находим на островах, даже таких больших, как Австралия или Южная Америка (которая долго была островом), весьма примитивные формы животных и растений.
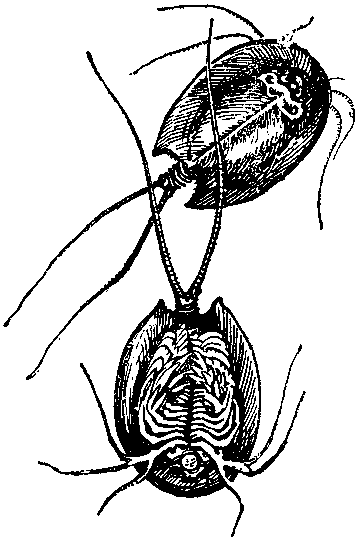
«Живое ископаемое» — рачок щитень, которого можно встретить в весенних лужах.
Для того чтобы познакомиться с «живым ископаемым», не обязательно ездить в Австралию. В весенних лужах под Москвой можно увидеть довольно крупного рачка щитня. От его облика веет какой-то подчеркнутой архаичностью, и это не удивительно. Щитень древнее многих динозавров, он существует с триасового периода без изменений, именно потому он обитает в пересыхающих, временных водоемах, где кроме него никто не может жить, разве только личинки двукрылых. Практически не имея врагов и конкурентов и обладая такой завидной адаптацией, как яйца, покрытые прочной оболочкой, переживающие сухой сезон в земле, щитень не «испытывал» никакой «потребности» к прогрессу.
Еще Дарвин указывал, что пресные, материковые воды — убежище многих архаичных организмов — «живых ископаемых». Причина этого в первую очередь — обедненность водных биоценозов — сообществ животных и растений.
В этом правиле есть, как будто бы, противоречие. Все мы знаем, хотя бы из описания, как исключительно богата флора и фауна тропиков, особенно тропических лесов. Казалось бы, вот где богатство биоценотических связей, наиболее сложные биоценозы! И однако мы находим в тропиках древние формы, давно вымершие в умеренной зоне. Оттуда описаны невзрачные растеньица, во многом аналогичные псилофитам — первым растениям, вышедшим на сушу в кембрийском периоде, а также саговники, гинкго и древовидные папоротники, под сенью которых в свое время прогуливались динозавры; самые примитивные пресмыкающиеся (крокодилы и гаттерия); примитивные лемуры и полуобезьяны — список этот можно увеличить.
Это кажущееся противоречие разрешил Шмальгаузен, указав, что каждое из «живых ископаемых» занимает весьма узкую экологическую нишу с весьма стабильными условиями, обладая к тому же каким-либо уникальным приспособлением, которое позволяет ему с успехом противостоять высшим, прогрессивным формам. Это специализация, благодаря которой «живые ископаемые» и могут процветать в узкоспециализированных условиях, подобно тому, как холодный сапожник выдерживает конкуренцию с мощными комбинатами бытового обслуживания.
Но среда может упрощаться и дальше. Выпадает внешний фактор — и отпадает нужда в органе. Что из этого следует? Среда уже не ведет отбор по развитию этого органа. В геноме начинают накапливаться неконтролируемые средой мутации, ведущие к дисгармонии развития, недоразвитию органа и, наконец, к полной его редукции.
Так потеряли способность к полету (а порой и сами крылья) многие птицы на островах без хищников, глаза — пещерные и подземные животные. Китообразные в начале третичного периода перешли в море, где органом движения у них стал мощный хвост. В таких условиях задние конечности не нужны, даже вредны, так как увеличивают сопротивление воды. Итогом явилась потеря задних конечностей, «ног». Передние остались, но приобрели функцию рулей. Тем не менее, в среднем каждый десятитысячный кашалот рождается с зачаточными задними ногами. А это может означать одно — в генотипе китообразных еще остались гены, ответственные за построение задних конечностей. Поэтому следует очень осторожно относиться к утверждению, что такая-то ветвь — тупиковая, она не сможет приспособиться к новой среде и ничего нового не даст. Чтобы установить наличие тупика, нужно уткнуться в стену — а стена где-то в отдаленном будущем.
Единственное, что можно утверждать: в настоящее время, в данных конкретных условиях эти группы, или же (что вернее) такие-то органы у них регрессируют, а сами они находятся в стадии морфофизиологического регресса, что, впрочем, не исключает их биологического прогресса, процветания.