Другой исследователь, А. Адриаансе, в той же Голландии, изучал тот же вид и заметил расхождение между своими данными и данными Бэрендсов — у него осы засыпали норку материалом, полученным при ее постройке и до этого сложенным аккуратной кучкой. У Бэрендсов аммофилы были не столь аккуратны и пользовались для этой цели любыми песчинками из окрестностей гнезда. Казалось бы, мелочь, пустяк — но Адриаансе, заинтересовавшись этим, открыл еще немало различий — его осы парализовали не гусениц, а личинок перепончатокрылых — пилильщиков (они очень похожи, их часто называют ложногусеницами). Нашлись и отличия в сроках размножения и ряде других деталей. Систематик Я. Вильке, узнав об этом, вспомнил, что экземпляры аммофил в музейных коллекциях различались, правда, отличия были ничтожными и их никто не считал видовыми. После сообщения Вильке Адриаансе установил, что эти две формы могут обитать совместно, на одном песчаном склоне, но никогда не скрещиваются. Пришлось создать новый вид — аммофилу Адриаансе. Вообще с видами-двойниками бывает, как с близнецами: познакомившись с ними, считаешь их неразличимыми, потом, наблюдая за ними изо дня в день, видишь, что один разговорчив, а другой молчаливее и, наконец, находишь «морфологические различия» — бородавку, родинку за ухом, лишнюю морщину. За время раздельной эволюции видов-двойников в них накапливаются различия, так что они перестают быть неразличимыми в коллекциях.
В заключение скажем еще несколько слов о пресловутой репродуктивной изоляции, то есть нескрещиваемости популяций. Мы уже убедились, что механизмы ее бывают разными — от разницы в сроках размножения до бесплодия гибридов. Она может предварять акт видообразования и может завершать его. Наконец, она может возникать скачком (полиплоидизация или крупная хромосомная перестройка) — тогда на пути потока генов между частями прежде единого вида как бы возникает плотина. Но, пожалуй, еще более часты случаи, когда поток генов превращается как бы в пересыхающий ручеек. Он исчезает постепенно, и главную роль в этом процессе играет дизруптивный отбор, отсеивающий менее приспособленные гетерозиготы. Вслед за Дарвином мы приходим к выводу, что видовые границы могут создаваться постепенно и на каком-то этапе оказываются условными.
Многих подобный вывод не устраивает; такие исследователи упорно ищут некий абсолютный критерий вида. Однако это часто напоминает поиски конца у кольца.
Изоляция и видообразование
Ретроспективный взгляд на историю эволюционного учения позволяет прийти к любопытному выводу. Авторы многих ошибочных взглядов исходили из в общем-то правильных положений. Ошибка заключалась в том, что эти положения абсолютизировались, роль каких-либо факторов преувеличивалась.
Так, Вейсман был совершенно прав, когда говорил о высокой стабильности наследственных задатков и о большей роли генетических рекомбинаций в эволюции. Действительно, эволюция реже использует новые гены, чем перетасовывает старые. Но считать гены полностью неизменяемыми — это уже ошибка.
Был прав и Де Фриз, открывший скачкообразное изменение признаков — мутации. Но считать, что мутационным путем возникают сразу виды, также будет ошибочным. Даже межвидовые гибриды растений долго «пришлифовываются» отбором к среде.
Учитель Вавилова В. Бэтсон полагал, что новые признаки возникают вследствие выпадения из генотипа какого-либо гена («теория присутствия — отсутствия»). Мы знаем, что дрейф генов и отбор могут привести к выпадению из популяции гена. Но ограничивать этим эволюцию нельзя.
По всей вероятности, сейчас чрезмерно высоко оценивается эволюционное значение ограничения панмиксности, а проще — генетической изоляции, причем генетическую изоляцию часто считают синонимом географической. Ход рассуждений здесь примерно такой: в панмиксной популяции не может сформироваться более одного нового вида, так как скрещивание исключает возможность образования двух сбалансированных генофондов. Как и в случае Дженкина, упор делается на «заболачивающее влияние скрещивания», но речь идет уже не об отдельных признаках и генах, а об их комбинациях. Поэтому, согласно этой точке зрения, видообразование может идти только в разных частях ареала (так называемое аллопатрическое видообразование). Разделение первоначально панмиксной популяции на два вида (симпатрическое видообразование) исключено.
Идея эта отнюдь не нова. Еще при жизни Дарвина немецкий натуралист М. Вагнер утверждал, что географическая изоляция — необходимое условие видообразовательного процесса, однако создатель теории естественного отбора с ним не согласился: «…я ни в каком случае не могу согласиться с этим натуралистом в том, что миграция и изоляция — необходимые элементы в процессе образования новых видов». Убеждение Дарвина в могуществе отбора осталось незыблемым, и сейчас мы можем только удивляться его прозорливости.
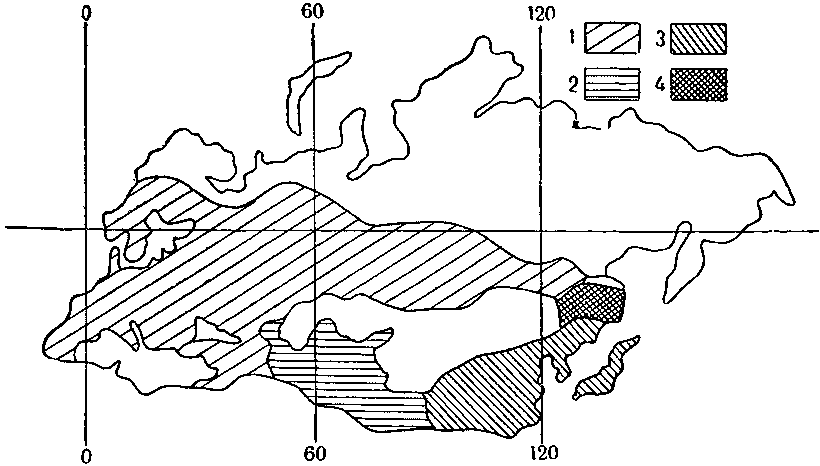
Распространение подвидов большой синицы: 1 — типичная форма; 2 — подвид бухарская; 3 — «большая малая»; 4 — зона, где синицы, распространявшиеся северным и южным путем, встречаются и не узнают друг друга.
Рассмотрим несколько примеров географической изоляции. Отлично изученная североамериканская леопардовая лягушка распространена очень широко — от Канады до Флориды. Соседние популяции ее хорошо скрещиваются, но самые северные и самые южные лягушки уже генетически изолированы и, по-видимому, достигли видового обособления, хотя провести границу между ними затруднительно. Она «размазана» по всей восточной части США.
Сходная картина наблюдается у многих видов птиц. Например, серебристая чайка и чайка-клуша в Западной Европе ведут себя как «хорошие» виды. Расселившись по побережью Северного Ледовитого океана, они замкнули в Европе кольцо распространения — и не узнали друг друга. Всем известная большая синица распространялась на восток двумя путями: северным — через Сибирь и Забайкалье и южным — через Среднюю Азию, Индию и Китай (пустынные нагорья Центральной Азии синиц не привлекали). Южная синица в пути несколько изменилась, она стала меньше, и этому подвиду присвоили довольно нелепое название — синица большая малая. Встретились они в Дальневосточном крае; здесь они не узнают друг друга, не скрещиваются. Все это хорошие примеры географического видообразования, аллопатрии. Несомненно, аллопатрические виды возникают в природе часто. Но современные последователи Вагнера полагают, что виды порождаются только таким путем.
Под категорию географического видообразования подводят возникновение видов из популяций предкового вида, обитающих практически совместно (например, мешетчатые крысы, одни из которых обитают на темных, а другие — на светлых почвах). Столь широкое трактование географии, конечно, не оставляет места для симпатрического видообразования (как бы близко ни обитали предковые особи, они неизбежно должны занимать разные точки пространства). Но это уже то, что математики называют приведением к абсурду.
Вспомним, однако, что существуют иные формы изоляции, кроме пространственной. Четвериков знал о них. Может ли такая изоляция возникнуть в пределах одной популяции?
Наиболее простые примеры экологической изоляции — это приспособление одного вида паразита к разным видам хозяев, что приводит в конце концов к радиации прежде единого вида на несколько видов, из которых каждый может обитать в одном лишь хозяине. Свиная и человеческая аскариды внешне морфологически неотличимы, однако яйца свиной аскариды в человеке не приживаются. Ясно, что они возникли от одного вида. Но когда произошло разделение? Быть может, пути человеческой и свиной аскарид разошлись, когда Моисей запретил правоверным иудеям употреблять в пищу свинину. Этот запрет был подтвержден Магометом — и в результате возникла изоляция между паразитами человека и свиньи. Если это так, то аскарида — пример вида, созданного религиозными запретами. Неисповедимы пути эволюции!