По Н.И. Волкову при рассмотрении факторов работоспособности мышц, в зависимости от основного механизма энергообеспечения, следует различать аэробную (окисление) и анаэробную работоспособность, а анаэробная работоспособность, в свою очередь, делится на лактатную (гликолиз) и алактатную (креатинфосфат). В качестве главных критериев оценки механизмов энергообеспечения мышечной деятельности принято выделять максимальную мощность, время удержания максимальной мощности, и общую емкость механизма. Максимальная мощность – это наибольшая скорость образования АТФ в данном метаболическом процессе. От мощности механизма энергообеспечения зависит возможная сила сокращения мышц в данном режиме работы. Под емкостью понимается общее количество энергии, которое можно получить за счет данного механизма ресинтеза АТФ.
Алактатная работоспособность мышц
Максимальная алактатная мощность, с одной стороны, зависит от концентрации и активности фермента креатинкиназа (переносящего фосфатную группу с креатинфосфата на АДФ) и собственно креатинфосфата, с другой стороны мощность данной реакции зависит от потребности мышц в энергии, соответственно, определяется максимальной скоростью расхода АТФ развиваемой мышцами. Максимальная длительность удержания алактатной мощности составляет 6-12 секунд. Алактатная емкость зависит от запасов креатинфосфата в мышце. О методах тренировки алактатной мощности и емкости я уже рассказывал ранее, рассматривая методы развития максимальной силы, и сейчас не буду подробно останавливаться на этом вопросе.
Лактатная работоспособность мышц
Максимальная лактатная мощность определяется главным образом концентрацией и активностью ключевых ферментов гликолиза. Время удержания максимальной мощности данного метаболического процесса составляет 30–60 секунд, и определяется, с одной стороны, устойчивостью ферментов гликолиза к понижению рН среды (повышение кислотности среды ингибирует активность гликолитических ферментов, что подавляет энергопроизводство), и устойчивостью кислотно-щелочного равновесия внутренней среды мышц, в условиях усиленной выработки лактата. С другой стороны, время удержания максимальной гликолитической мощности лимитируется факторами утомления мышцы, снижающими интенсивность сокращения.
Из вышесказанного следует, что для запуска адаптационных процессов, направленных на увеличение максимальной гликолитической мощности, длительность нагрузки должна соответствовать времени удержания максимальной мощности данного метаболического процесса, что составляет 30–60 секунд. Отдых между подходами должен быть достаточно длительным, для обеспечения вывода продуктов метаболизма из мышцы и развития высокой мощности гликолиза в следующем подходе. Устойчивость рН среды мышечных волокон к выбросу молочной кислоты и устойчивость ключевых ферментов к снижению рН вырабатывается в ходе тренировок, сопровождающихся максимальным накоплением лактата в мышцах. Это могут быть нагрузки высокой интенсивности, длительностью 1–1.5 минуты до наступления отказа мышц, вызванного сильным закислением, либо более короткие нагрузки, длительностью 20–40 секунд, со столь же коротким интервалом отдыха, приводящие к кумулятивному накоплению лактата в мышцах.
Гликолитическая емкость определяется главным образом запасами гликогена в мышцах, гликоген печени для процессов гликолиза не обладает достаточной мобильностью. О методах накопления мышечного гликогена, как и гликогена печени, я уже рассказывал при рассмотрении факторов общей работоспособности организма.
Аэробная работоспособность мышц
Максимальная аэробная мощность зависит главным образом от плотности митохондрий в мышечных волокнах, концентрации и активности окислительных ферментов, скорости поступления кислорода вглубь волокна. Объем кислорода доступного для окислительных реакций лимитируется, как факторами общей работоспособности организма, которые я уже ранее рассматривал, так и рядом локальных внутримышечных факторов, среди которых можно выделить капиляризацию мышц, концентрацию миоглобина, диаметр мышечного волокна (чем меньше диаметр волокна, тем лучше оно снабжается кислородом и тем выше его относительная аэробная мощность). Скорость производства АТФ за счет окисления достигает максимальных значений на 2-3-й минуте работы, что связано с необходимостью развертывания множества процессов, обеспечивающих доставку кислорода к митохондриям. Время удержания максимальной аэробной мощности составляет примерно 6 минут, в дальнейшем аэробная мощность снижается по причине усталости всех активно работающих систем организма. Соответственно, для повышения аэробной мощности мышц тренировочная нагрузка должна длиться не менее 2 минут (для выхода скорости энергопроизводства на максимум). Не имеет смысла и затягивать нагрузку дольше чем на 6 минут, при тренировке именно мощности, так как далее идет ее (мощности) снижение. Эффективным оказывается многократное повторение таких нагрузок.
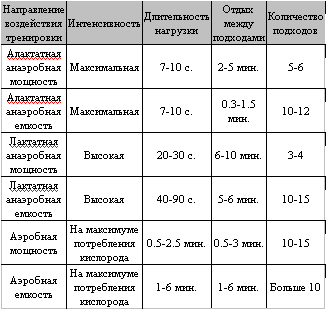
В заключение хочу привести сводную таблицу тренировочного воздействия на работоспособность мышц в различных режимах работы, почерпнутую мной из диссертации М. Хосни, посвященной изучению биохимических основ интервальной тренировки. Для развития соответствующих качеств Хосни рекомендует следующие методические приемы:
На этом я заканчиваю изложение основ тренировки работоспособности мышц и перехожу к анализу основных факторов, определяющих мышечные объемы спортсмена.
Часть 6
Рассмотрение тренировочных методик
Ну что же, основные методы тренировок, способствующих развитию силы и силовой выносливости мышц, нами уже рассмотрены. Настало время приступить к рассмотрению тренировочных методик, в полной мере способствующих гипертрофии мышц, для чего следует определить тканевые и внутриклеточные структуры, от развития которых зависят мышечные объемы спортсмена. Я уже затрагивал немного этот вопрос во второй части, теперь остановимся на нем чуть подробнее. Как вы помните, объем мышцы, прежде всего, определяется количеством мышечных волокон (клеток) в теле мышцы, размером самих этих волокон, а так же объемом межклеточного вещества, представленного, главным образом, кровеносными сосудами и соединительной тканью, отделяющей друг от друга отдельные волокна и их пучки. Немаловажное значение для визуальных объемов спортсмена имеют и запасы жира в организме, однако, вклад жиров уже трудно назвать вкладом в «мышечные» объемы, а стандарты соревновательного бодибилдинга требует минимизации такового вклада, поэтому методы тренировок, приводящих к увеличению жировой составляющей объемов спортсмена, я рассматривать не буду, они и так хорошо всем известны. Увеличение количества мышечных волокон у человека ни разу достоверно не фиксировалось в экспериментах, хотя, как я уже говорил ранее, гиперплазия не кажется мне столь уж невероятным явлением, после того так она была зафиксирована у животных, но, дабы не прослыть пустым фантазером, я не буду включать гиперплазию в причины мышечной гипертрофии, до появления достоверных экспериментов, зафиксировавших увеличение количества мышечных волокон у человека. И так, нам остается уповать только на капилляризацию мышц, увеличение в объеме мышечных волокон и рост соединительной ткани. Объем мышечных волокон контролируется, прежде всего, количеством мышечных ядер в волокне. Именно от количества ядер, при прочих равных условиях, зависит общий объем белка, синтезируемого мышечным волокном в единицу времени. И этот фактор незаслуженно игнорируется многими специалистами при рассмотрении причин мышечной гипертрофии под действием тренировок. Как вы помните, к увеличению количества мышечных ядер приводит деление клеток сателлит, инициируемое факторами, появляющимися в мышечном волокне при его повреждении. Но ядра служат первопричиной, а увеличивают объем волокна иные клеточные структуры, такие как миофибриллы, саркоплазма, митохондрии, и др. Вот какие данные о потенциале роста мышц, за счет различных клеточных и межклеточных структур приводит в Ф. Хетфилд: