Исключение составляют скаковые лошади и некоторые породы собак, при выведении которых человек вел отбор на максимальную скорость бега. В этом случае цели человека не слишком расходились с обычными «целями» естественного отбора. В итоге деградация лошадей (прежде всего, уменьшение их размеров) связана обратной зависимостью с уровнем развития государства и его военных институтов. В частности, с падением Рима начался процесс резкого измельчания лошадей.
Попробуем просуммировать некоторые особенности, отличающие домашних млекопитающих от их диких предков. Они включают в себя изменение характера окраски шкуры (прежде всего многочисленные пегости); потерю или редукцию рогов; уменьшение числа хвостовых позвонков; уменьшение размеров тела и уменьшение величины головного мозга и органов, связанных с восприятием внешних сигналов; вислоухость (т. е. неспособность двигать ушами). Все эти изменения свидетельствуют о деградации функций, повышающих выживание в естественной (дикой) обстановке. Пожалуй, только функция размножения была усилена в ходе доместикации, что и не удивительно, так как, стремясь увеличить продуктивность животных, человек автоматически отбирал особей, приносящих наибольшее число потомков.
Итак, сходный (конвергентный) характер морфологических изменений при доместикации эволюционно неродственных видов скорее всего обусловлен снижением давления очищающего отбора, чему в немалой степени мог способствовать и интенсивный искусственный отбор, направленный на улучшение хозяйственно ценных признаков. Однако существует еще одно объяснение этой конвергенции. Советский генетик Д.К. Беляев предположил, что главная роль в процессе одомашнивания принадлежит отбору животных на ручной (неагрессивный) тип поведения. В ходе долговременного отбора на ручной тип поведения у серебристо-черных лисиц Д.К. Беляев с сотрудниками обнаружили немало наследуемых морфологических отклонений, вполне типичных для процесса одомашнивания.
По мнению ученого, главная особенность отбора на поведение состоит в том, что он ведет к изменению гормонального статуса животных, а это, в свою очередь, может явиться причиной глубокой и направленной реорганизации онтогенеза. Постулируется, что изменение уровня гормонов может привести к наследуемому изменению активности ряда генов. Набор этих генов не вполне случайный, что и объясняет параллелизм в появлении аномальных форм у разных видов в процессе доместикации. Заметим, однако, что данная концепция оставляет без ответа многие вопросы. Получается, что сдвиг гормонального статуса взрослого организма может вызвать направленное изменение активности вполне определенных генов в клетках зародышевого пути, причем это измененное состояние генов способно сохраняться у потомков. Тем самым фактически предполагается превращение средовой (модификационной) изменчивости в наследственную.
Отбор и интеграция процесса развития
В ходе искусственного отбора происходит глубокая перестройка генофонда популяции, поэтому не следует думать, что из всех предложенных объяснений сопротивления популяции отбору верно только одно. Хотя мы и далеки от мысли, что все существующее разумно, но если природная популяция не гибнет в течение тысяч поколений, то, по-видимому, организация ее генофонда не так уж далека от оптимальной.
Популяция не в силах полностью оградить себя от таких факторов, как мутационное давление, генный дрейф или генный поток, поэтому она всегда отягощена различными формами генетического груза. В то же время постоянное действие естественного отбора направлено на сохранение максимального в данных условиях уровня приспособленности. Для достижения этой цели популяция использует все доступные ей средства: от изменения способа размножения и уровня кроссоверной рекомбинации до изменения схемы индивидуального развития.
Выше мы только вскользь упомянули о гипотезе С. Райта, которая объясняет сопротивление популяции действию отбора тонкой «подогнанностью» друг к другу аллелей разных локусов. Такая взаимозависимость эффектов генов скорее всего должна проявляться в процессе индивидуального развития организма.
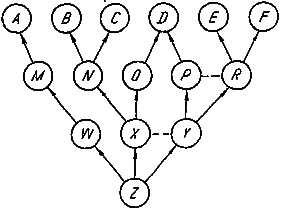
Рассмотрим абстрактную схему формирования взрослых структур А — F (рис. 15). За каждой из них стоит ряд зародышевых структур, нередко взаимодействующих друг с другом. На схеме эти зачатки условно разделены на два класса: поздние М — Р и ранние W— Y. Формально зиготу тоже можно считать зачатком (Z), общим для всех взрослых и эмбриональных структур организма. Каждая сплошная стрелка на схеме олицетворяет собой все многообразие генов, ответственных за развитие более раннего зачатка в более поздний. Эти гены можно символизировать сочетанием двух прописных букв — первая относится к более раннему зачатку, вторая — к более позднему. Взаимодействия типа эмбриональной индукции обозначены на схеме горизонтальными пунктирными линиями.
Представим себе, что селекционер стремится изменить вполне конкретную взрослую структуру. Ясно, что для этого он должен в каких-то генах, скрытых за стрелками, ведущими к данной структуре, зафиксировать новые (благоприятные с точки зрения селекционера) аллели. Из рис. 15 видно, что для изменения структуры А можно замещать аллели в генах ZW, WM и МА.
Рис. 15. Схема индивидуального развития структур организма.
Z — зигота; W—Y — ранние зачатки; M — R — поздние зачатки; А — F — взрослые структуры. Штриховой линией отмечена эмбриональная индукция.
Обратим внимание на путь формирования структуры В. Отметим, что из позднего зачатка N кроме нее возникает еще одна взрослая структура С. Если эта последняя прямо связана с основными показателями приспособленности, то ее развитие должно быть оптимизировано естественным отбором. Для изменения структуры В можно замещать аллели в любых локусах, стоящих за стрелками, ведущими к ней. Однако такие замещения в ранних генах кроме изменения структуры В могут затронуть еще одну структуру — С, а это уже может снизить уровень главных показателей приспособленности. Чтобы скомпенсировать данный негативный эффект, следовало бы структуре С возвратить ее прежнее значение, зафиксировав для этого новый аллель по одному из поздних локусов NC, т. е. в данном случае для адаптивного сдвига нужно зафиксировать два новых аллеля.
Обобщая этот пример, можно показать, что чем больше нитей связывает развитие какой-то структуры с другими, тем больше новых аллелей нужно зафиксировать для достижения одного и того же сдвига структуры у взрослой особи. Выходит, что отбор в основном должен оперировать с генами, которые управляют завершающими стадиями развития организма, тогда как гены, контролирующие ранние стадии, фактически не допускают изменений. Это хорошо объясняет сохранение основного плана строения животного в ходе эволюции. В то же время, чем раньше обособляется зачаток, тем выше шансы на эволюционный сдвиг у развивающейся из этого зачатка структуры.
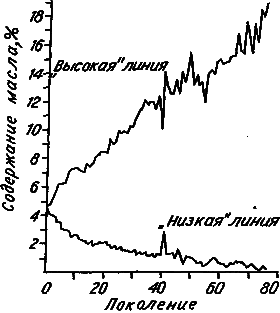
Любопытно, что у растений негативные эффекты искусственного отбора выражены гораздо меньше, чем у животных. Особенно впечатляюще выглядят результаты знаменитого Иллинойсского эксперимента. Группа американских селекционеров в самом конце XIX века приступила к отбору, направленному на изменение ряда количественных признаков у кукурузы. Этот опыт продолжается до сих пор. На рис. 16 показано, как изменялось в ходе 76 поколений отбора содержание масла в семенах. Как видно, ответ на отбор может сохраняться в течение весьма продолжительного периода.
Рис. 16. Отбор на изменение содержания масла в семенах кукурузы (по: [Dudley, 1977]).
Эту особенность растений можно объяснить с двух позиций. Во-первых, в отличие от животных в жизненном цикле растений есть стадия гаплоидного организма — гаметофита. Всем известная цветочная пыльца является скоплением мужских гаметофитов. Биомасса пыльцевого зерна — продукт деятельности гаплоидного генома одного ядра. Этот же геном обеспечивает превращение пыльцевого зерна в трубку, прорастающую сквозь пестик до завязи. По этой трубке в зародышевый мешок доставляются мужские гаметы. Избыток пыльцевых зерен создает условия для соревнования гаметофитов по оплодотворяющей способности, т. е. на стадии мужского гаметофита идет как бы проверка большого числа генов на качество их работы. Это можно рассматривать как приспособление для снижения мутационного генетического груза.