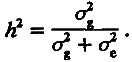S = Mc — M. (4.16)
Аналогично можно ввести представление об ответе популяции на отбор (R) как о сдвиге среднего значения популяции за одно поколение отбора:
R = M'c — M, (4.18)
где M'c — среднее значение признака у потомков «счастливцев».
Большой экспериментальный материал свидетельствует, что ответ на отбор прямо пропорционален селекционному дифференциалу. Следовательно, чем сильнее давление отбора, тем больше величина ответа на него. Величина этого ответа — внутреннее (генетически обусловленное) свойство популяции. Его количественной мерой является наследуемость — коэффициент пропорциональности (h2), связывающий ответ на отбор с селекционным дифференциалом. Итак, при ответе конкретной популяции на отбор по конкретному количественному признаку выполняется равенство
R = h2S. (4.19)
Наследуемость разных признаков у особей одной популяции может существенно различаться. Кроме того, может различаться и наследуемость одного и того же признака в разных популяциях одного вида. Что же лежит в основе этого явления?
Пусть мы имеем дело с популяцией, состоящей из генетически идентичных особей. Многократно показано, что при любом давлении отбора среднее значение любого признака в следующем поколении практически не изменяется (h2 = 0). Впервые это было продемонстрировано в знаменитом эксперименте В. Иогансена на чистых линиях фасоли. Иными словами, наследуемость равна нулю, если равна нулю генетическая компонента изменчивости.
Представим на момент, что мы в состоянии по фенотипу особи опознать ее генотип. Если к тому же эти особи размножаются бесполым путем, то состав генотипов потомков будет совпадать с таковым у отобранной группы родителей. Следовательно, и средние значения признаков потомков будут совпадать с их средними значениями у родителей, тем самым величина ответа на отбор сравняется с селекционным дифференциалом. Очевидно, что в данном случае h2 = 1.
Обычно же ответ на отбор меньше селекционного дифференциала, так как часть особей попадает в отобранную группу совершенно случайно (вследствие счастливого стечения средовых факторов), поэтому особенность их фенотипа наследоваться не может. Ведь для того, чтобы потомки этих «баловней судьбы» (а не генотипа) попали в отбираемую группу, им снова должно повезти. Но случай на то и случай, чтобы не повторяться регулярно.
В генетике количественных признаков доказывается, что наследуемость равна доле генетической дисперсии в полной фенотипической дисперсии признака. Вспоминая, что эта последняя складывается из генетической и средовой компонент, сказанное можно передать формулой
(4.20)
Повторяя процедуру отбора, направленного на изменение величины признака, в течение большого числа поколений мы можем в конце концов добиться фиксации генотипов, включающих в себя только сильные или только слабые аллели (в зависимости от направления отбора). Ясно, что в таком случае весь запас генетической изменчивости будет исчерпан, и дальнейший ответ на отбор станет невозможным.
Теперь обратимся к эксперименту. Действительно, у животных через несколько поколений отбора ответ на него обычно прекращается, и среднее значение признака выходит на плато. Однако интерпретировать это явление не так просто.
Негативные эффекты искусственного отбора
Первый лабораторный эксперимент по отбору на изменение количественных признаков был проведен в начале XX века американским генетиком У. Кастлом. Он ставил перед собой цель — превратить пегих (пятнистых) крыс в одноцветных. В одном варианте отбор был направлен на увеличение площади белых пятен, в другом — в противоположном направлении. Примерно через 20 поколений Кастл был близок к цели, т. е. ему удалось получить почти белых и почти черных крыс, но опыт пришлось прекратить, так как животные захирели и перестали размножаться. Короче говоря, у крыс резко снизились основные показатели приспособленности.
В дальнейшем сходные результаты были получены на большом числе животных объектов — мышах, курах, золотых рыбках, мучных хрущаках и, конечно, на дрозофиле. Везде по мере сдвига самых разных количественных признаков в любом направлении наблюдалось снижение показателей приспособленности: прежде всего жизнеспособности и плодовитости. Популяции как бы сопротивлялись действию искусственного отбора.
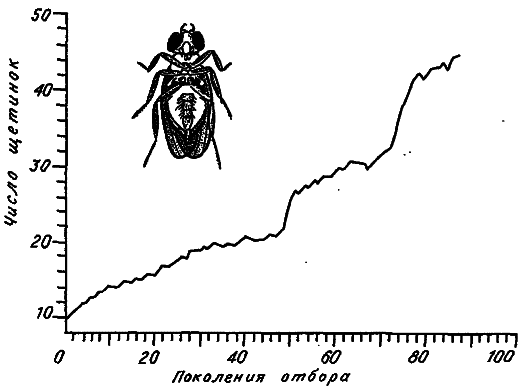
Особенно много экспериментов проведено по увеличению числа щетинок на разных частях тела у дрозофилы. Пристрастие именно к этому объекту объясняется очень коротким жизненным циклом плодовой мушки и ее хорошей генетической изученностью. Пример такой селекции приведен на рис. 14. Величина признака сначала монотонно возрастает, затем (обычно после 15–20 поколений отбора) наступает застой, ответ на отбор прекращается, а жизнеспособность и плодовитость мух падают. Оказывается, если отбор прекратить, то основные показатели приспособленности довольно быстро восстанавливаются, однако обычно это сочетается с существенным снижением числа щетинок, т. е. за несколько поколений «отдыха» от отбора величина признака сдвигается назад в сторону его исходных значений. Если после периода «отдыха» отбор возобновить, то можно снова поднять признак на прежний уровень, иногда даже при более высоких значениях главных компонент приспособленности.
Для экспериментов по долговременному отбору на увеличение числа щетинок характерна поразительная невоспроизводимость динамики изменения признака. В то же время в этой картине при всем ее многообразии имеются некоторые общие моменты. Обычно все начинается с довольно быстрого ответа на отбор, потом скорость увеличения признака постепенно падает, и кривая отбора выходит на плато. В этом состоянии «застоя», несмотря на продолжающуюся селекцию, популяция может находиться неопределенно долго. Затем внезапно может наступить фаза быстрого ответа на отбор, и кривая выходит на новое, более высокое плато. Этап нового застоя через какое-то число поколений может опять смениться фазой внезапного ответа, потом снова наступает застой и т. д.
Рис. 14. Отбор на увеличение числа абдоминальных щетинок у дрозофилы Drosophila melanogaster (по: [Yoo, 1980]).
Вся эта феноменология выглядит весьма загадочно и до сих пор не получила удовлетворительного объяснения. Существует несколько гипотез. Самая простая из них связывает падение приспособленности с так называемой инбредной депрессией. Действительно, обычно численность лабораторных понуляций невелика и в каждом поколении большая их часть подвергается браковке. Поэтому с ходом отбора постепенно повышается степень родства особей, вступающих в скрещивание. А это означает прогрессивное возрастание степени гомозиготности генов, часть которых может быть представлена вредными рецессивными аллелями. Однако расчеты показывают, что инбредная депрессия повинна лишь в части негативных последствий селекции.
Вторая гипотеза, высказанная выдающимся американским генетиком и математиком С. Райтом, объясняет негативный эффект селекции нарушением взаимодействия генов, хорошо «подогнанных» друг к другу естественным отбором. В ходе искусственного отбора в генофонде популяции нарастает концентрация аллелей, которые сдвигают признак в направлении, нужном экспериментатору, но плохо «сочетаются» с аллелями других генов.
Третья гипотеза принадлежит знаменитому английскому генетику К. Мазеру. Он предположил, что степень развития признака определяется совокупным эффектом большого числа локусов, аллели которых могут сдвигать величину признака в любом направлении. По мнению Мазера, слабые и сильные аллели (по эффекту на признак) разбросаны вдоль хромосомы случайно, а между ними находятся гены, влияющие на приспособленность и которые также представлены своими сильными и слабыми аллелями.