Теперь рассмотрим совокупность всевозможных финалов частоты аллеля А1. У части финальных популяций она будет равна единице, а у всех других — нулю. Очевидно, что доля популяций, зафиксировавших в конце концов аллель А1 (иначе говоря, вероятность фиксации данного аллеля), равна его средней частоте в полной совокупности финальных популяций. Поскольку (см. формулу (3.25)) последняя должна равняться частоте аллеля в исходной популяции (р0), мы приходим к нетривиальному заключению: вероятность фиксации нейтрального аллеля равна его частоте в генофонде популяции. Например, если эта частота равна 0,5, то аллель с равной вероятностью может или исчезнуть из популяции, или в ней зафиксироваться. Теперь перейдем к очень важному следствию — к судьбе только что возникшей мутации.
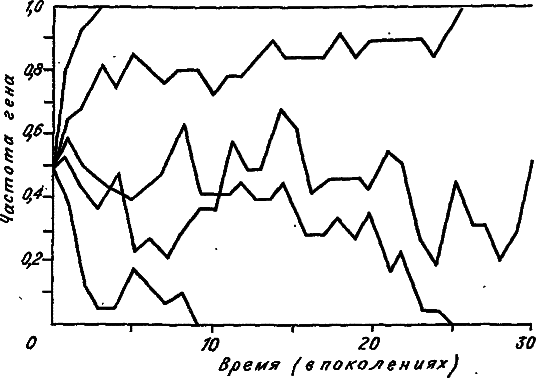
Рис. 11. Дрейф аллельных частот в пяти машинных экспериментах. Размер популяции N = 10. Исходная частота обоих аллелей 0,5 (по: [Kimura, 1983]).
Пусть в популяции возник совершенно новый нейтральный аллель. Очевидно, что его частота в момент возникновения равна 1/2N, поэтому и зафиксироваться с помощью дрейфа он может с той же вероятностью. Этот результат приводит нас к грустному выводу: фактически любой вновь возникший аллель обречен на исчезновение, поскольку вероятность этого события чрезвычайно близка к единице (1 — 1/2N). В то же время заметим, что процесс мутирования постоянно ведет к появлению новых аллелей. Если через V обозначить мутабильность локуса, то в каждом поколении в популяции должно появляться 2NV мутантных аллелей. Поскольку они не различаются естественным отбором, любой из них может зафиксироваться с вероятностью 1/2N. Значит, в данном случае скорость фиксации любого мутантного аллеля только за счет генного дрейфа должна равняться 2NV(1/2N). Обозначив скорость фиксации мутантного аллеля через К, приходим к очень простому уравнению
K = V. (3.26)
Этот изящный результат, полученный М. Кимурой в 1960 г., показывает, что в популяциях (вне зависимости от их размеров) может идти процесс фиксации новых аллелей, скорость которого определяется только скоростью мутирования локусов. Однако среднее число поколений, нужное для фиксации мутантного аллеля с помощью дрейфа, пропорционально размеру популяции и, как показал Кимура, оно близко к 4N.
Заметим, что геном любого вида содержит несколько тысяч локусов, поэтому в генофонде любой популяции только вследствие неизбежного генного дрейфа почти в каждом поколении фиксируются какие-то аллели. Однако подчеркнем, что этот вывод мы получили при допущении, что жизнеспособность организма не зависит от аллельного состава локусов; иными словами, естественный отбор эти аллели не различает. Несмотря на то что фиксация нейтральных аллелей занимает много времени, у данного процесса есть одна выгодная для популяции особенность — он проходит для нее совершенно бескровно.
Движущий отбор
До сих пор мы имели дело с панмиктической популяцией конечного размера, занимающей ограниченную территорию с фиксированными условиями среды. На самом же деле мир, в котором обитает популяция, изменчив, и идеально приспособиться к нему невозможно. Представим себе, что наша популяция в какой-то момент времени хорошо приспособлена к условиям своего существования, однако вдруг эти условия начали медленно, но неуклонно ухудшаться. В данной ситуации перед популяцией встает проблема огромной сложности: нужно не просто отбраковать относительно редкие мутантные аллели (чему в немалой степени помогает генетический дрейф), но и заменить старые, далеко не дефектные аллели на иные, более благоприятные, т. е. лучше соответствующие новым условиям среды.
Будем считать, что благоприятные мутации полудоминантны, т. е. h = 0,5. Если принять приспособленность гомозигот по старому аллелю (w11) за единицу, то приспособленность гомозигот по благоприятному аллелю (w22) должна равняться (1+s), а гетерозигот — (1+0,5s). По аналогии с рассмотренным выше случаем вредных мутаций, заменяя — s на +s, можно рассчитать среднюю приспособленность популяции (w) и среднюю приспособленность доноров благоприятного аллеля (w2):
w = 1 + qs, (3.27)
w2 = 1 + 0,5ps + qs, (3.28)
тогда сдвиг частоты благоприятного аллеля за одну генерацию можно определить по формуле
Δq = (0,5pqs) / (1 + qs). (3.29)
При малых значениях q и s знаменатель в правой части этого уравнения мало отличается от единицы, поэтому с хорошей точностью должно выполняться соотношение
Δq ≈ 0,5pqs, (3.30)
т. е. скорость возрастания частоты благоприятного аллеля в генофонде популяции прямо пропорциональна его частоте и селективному преимуществу.
Глядя на соотношение (3.30), можно подумать, что были бы в наличии благоприятные мутации, а процесс их фиксации — дело десятое. Однако анализ решения этого уравнения показывает, что даже при таком высоком коэффициенте отбора, как 0,1, для достижения частот благоприятного аллеля, близких к единице, требуется около 100 поколений, при s = 0,01 — не менее 1000.
В этих оценках мы не учитывали ограниченность размера реальных популяций. Кроме того, мы полагали, что каждый носитель благоприятной мутации имеет все возможности реализовать свое селективное преимущество. На самом же деле, разбиение вида на небольшие популяции является весьма эффективным средством не столько для фиксации, сколько для удаления редких аллелей — как нейтральных, так и благоприятных.
Для того чтобы новый благоприятный аллель вытеснил старый, нужно еще и простое везение. К сожалению, мы сейчас убедимся, что шансы на такое везение не очень велики. Представим себе, что в популяции сколь угодно большого, но постоянного размера возник благоприятный аллель А2. Его носителем может быть только гетерозигота А1А2. Поскольку размер популяции постоянный, каждая особь передает в генофонд следующего поколения в среднем только две копии локуса А. Значит, гетерозигота А1А2 может передать эти две копии в виде одной из трех комбинаций: А1А1, А2А2 и А1А2. Первая комбинация означает потерю аллеля А2, вероятность этого события равна 25 %. В 50 % случаев в генофонд следующего поколения будет передана только одна копия мутантного аллеля (и в следующем поколении он может снова с вероятностью 25 % потеряться), а в 25 % случаев количество мутантных аллелей удвоится.
В этом рассуждении мы не учитывали эффекта статистического варьирования числа потомков каждого родителя. Точное решение задачи было получено выдающимся английским математиком и генетиком Р. Фишером. Он продемонстрировал, что вероятность потери вновь возникшей нейтральной мутации в первом же поколении равна 0,37 и быстро растет с увеличением числа поколений, стремясь в конечном счете к единице. Если же мутантный аллель сообщает его носителю селективное преимущество s в гомозиготном состоянии и 0,5s в гетерозиготном, то вероятность его фиксации численно близка к s. Итак, если коэффициент отбора в пользу носителя одной копии благоприятного аллеля составляет целых 5 %, то все-таки шансы его утратить равны 90 %. Следует заметить, что обычно s намного меньше единицы и даже меньше 0,1.
Положение может спасти лишь повторное возникновение благоприятных мутаций. Если скорость их возникновения равна v, то в популяции с размером N за одно поколение должно появляться 2Nv новых благоприятных мутаций, из которых будет фиксироваться доля, численно равная s. Отсюда скорость фиксации полудоминантных благоприятных мутаций (K) определяется уравнением