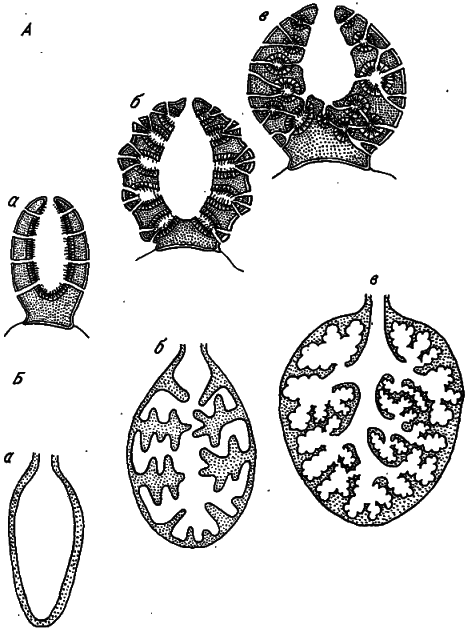
Тот же тип геометрического усложнения можно обнаружить и у самых примитивных животных — губок. Здесь традиционно выделяют три ступени прогрессивного развития — аскон, сикон и лейкон (рис. 4, А). Усложнение сопряжено с увеличением доли клеток, добывающих пищу, — хоаноцитов.
Рис. 4. Усложнение двухмерных структур (по: [Хадорн, Венер, 1989]).
А — усложнение губок: а — аксон, б — сикон, в — лейкон; Б — усложнение легких у позвоночных: а — хвостатые амфибии, б — рептилии, в — млекопитающие.
Можно выделить следующие типы геометрического усложнения. Для одномерных структур — линий, стержней, тонких трубок и балок — это иерархия изгибов и ветвлений, для двухмерных — иерархия складок, выпячиваний, карманов. В обоих случаях повышается размерность. Одномерная лопастная линия своими изгибами может плотно покрыть двухмерную поверхность стенки раковины. Система одномерных клеточных балок создает трехмерную структуру печеночной паренхимы. То же можно сказать о системе тонких трубочек, укладываемых в спирали. Выпячивания двухмерных структур создают трехмерные структуры губчатого или слоистого строения. Для трехмерных структур усложнение часто сопряжено просто с повторением одних и тех же элементов (нефронов, семенных канальцев, мышечных волокон и т. д.). В итоге растет масса органа. Главный показатель, сопровождающий усложнение структуры, заключается в увеличении интенсивности ее функции. Если такое усложнение наблюдается в ходе исторического развития, то его принято называть «прогрессивным».
Усложнение в ходе онтогенеза
К проблеме структурной сложности можно подойти и с другой стороны. Обратим внимание на то, что любой многоклеточный организм развивается из одной, хотя и очень большой, клетки — оплодотворенного яйца. Эта клетка, если ее изучать под микроскопом, производит впечатление весьма однородного и бесструктурного объекта. И тем не менее гомогенная масса куриного желтка через 21 день насиживания превращается в цыпленка со всеми атрибутами сложного организма. Суть этого процесса, по мысли К. Бэра, сводится к следующему: «Во время развития из гомогенного и общего постепенно возникает гетерогенное и частное».
Еще натурфилософы говорили о параллелизме между индивидуальным развитием высших организмов и лестницей существ. Однако тот же Бэр показал, что на самом деле имеет место не повторение стадий, подобных ступенькам лестницы существ, а сходство начальных стадий развития у организмов одного типа (в смысле Ж. Кювье). У всех систематически близких видов можно обнаружить несколько стадий, когда зачатки гомологичных органов располагаются по отношению друг к другу весьма сходным образом, как бы по одному плану. Так, у всех зародышей позвоночных имеется внутренний скелет в виде продольного тяжа — хорды. Выше нее (дорсальнее) располагается нервная трубка, а ниже (вентральнее) — первичная кишка.
Образование первичных зародышевых структур у большинства хордовых протекает удивительно сходно. Возьмем, к примеру, нервную трубку — зачаток центральной нервной системы. У всех хордовых можно видеть, как в однородной эктодерме ранней гаструлы в области ее контакта с первичной кишкой постепенно обособляется узкая продольная полоска клеток — нервная пластинка. Затем она прогибается и, пройдя фазу желобка, замыкается в нервную трубку, отделившись от сросшейся над нею эктодермы.
Еще в 1828 г. Бэр пришел к заключению, что, во-первых, едва ли правомочно сопоставлять уровень организации животных, принадлежащих разным типам, и, во-вторых, у представителей одного типа этот уровень можно оценить по числу гистологических и морфологических дифференцировок, т. е. по числу деталей, выявляемых на глаз и под микроскопом при разном увеличении.
В основе морфологической дифференцировки лежит несколько фундаментальных процессов, протекающих на клеточном уровне: клеточное размножение, клеточное движение, межклеточное взаимодействие, клеточная дифференцировка и, наконец, клеточная гибель. Пожалуй, самым важным для нашей темы является процесс клеточной дифференцировки.
У многоклеточных организмов каждая клетка находится в одном из целого ряда весьма устойчивых дискретных состояний — клеточных типов, или дифференцировок. Клетки разных типов отличаются по огромному числу свойств: скорости размножения, спектру синтезируемых молекул, сродству к другим клеткам или внеклеточным структурам, способу клеточного движения и т. д. Одни из них стремятся агрегировать друг с другом, чтобы образовать шаровидные скопления или тонкие, в один клеточный слой, пласты. Другие, наоборот, как бы избегают контактов, третьи тяготеют к бесклеточным мембранам, а четвертые пытаются соединиться с клетками других типов.
Самым важным здесь является момент изменения клеткой своего типа. Это событие может произойти по разным причинам. Клетка может дифференцироваться совершенно самостоятельно (возможно, израсходовав какой-то ресурс или наработав какой-то продукт). Данный путь обычен для самых начальных стадий индивидуального развития, в особенности у животных, которых принято помещать на нижние ветви филогенетического древа. Однако чаще всего клетка дифференцируется под влиянием внешнего сигнала, поступившего от других клеток того же зародыша. Этот тип дифференцировки, весьма характерный для позвоночных, получил название эмбриональной индукции.
В данном случае сигнал к изменению дифференцировки передается от клеток индуктора к близко расположенным клеткам ткани, отвечающей на индукцию. Клетки нового типа могут затем сами выступать в качестве индуктора, побуждая к дифференцировке другие клетки, иными словами, возникает целый каскад (иерархия) эмбриональных индукций. Необходимость довольно тесных контактов между обоими участниками индукционного процесса объясняет эволюционный консерватизм взаимного расположения многих эмбриональных зачатков, т. е. сохранение в эволюции основного плана строения зародышевых структур.
В связи с этим в качестве еще одной меры сложности организма можно выбрать число клеточных типов, встречающихся на всех стадиях его развития. К сожалению, тщательный подсчет данного числа пока не проведен, хотя ясно, что многообразие клеточных типов у насекомых и позвоночных измеряется сотнями, а у червей и других низших беспозвоночных — только десятками. Самая примитивная группа многоклеточных животных — губки — располагает едва ли одним десятком разных дифференцировок. Таким образом, наше интуитивное представление о повышении структурной сложности организмов в ходе исторического развития жизни на Земле, по-видимому, нас не обманывает.
Сериальная гомология
В строении многих животных, в том числе и у доминирующих в наше время позвоночных и членистоногих, можно обнаружить весьма сходные морфологические структуры, повторяющиеся вдоль продольной оси организма. Такие структуры называют метамерами, а само явление — сериальной гомологией. У позвоночных метамерами являются позвонки, ребра, сомиты и т. д. У кольчатых червей и членистоногих — это, прежде всего, сегменты.
У некоторых кольчатых червей (считающихся примитивными) все сегменты с их наружными придатками поразительно сходны. Лишь два из них — головной и конечный — составляют исключение. У других кольчецов (эволюционно «более продвинутых») можно выделить отделы с несколько иным строением сегментов — так называемые тагмы. У членистоногих дифференциация сегментов (тагмозис) зашла гораздо дальше, особенно у высших ракообразных и насекомых. Сравнивая представителей различных групп кольчатых червей и членистоногих, можно получить плавный ряд возрастающей дифференциации метамеров. При этом можно заметить, как простые наружные придатки сегментов кольчатых червей преобразуются у членистоногих в конечности, состоящие из набора трубчатых члеников. Строение конечностей в разных тагмах может поразительно различаться, отражая их функциональную дифференциацию. Они могут быть ногами, органами чувств, хватательными и ротовыми придатками, органами дыхания, приспособлениями для спаривания и для защиты потомства (рис. 5).