Экклсу не пришлось долго ждать опровержения своей гипотезы. Вернувшись в Университетский колледж Лондона, Кац получил прямые доказательства того, что именно ацетилхолин, выделяемый мотонейронами, является причиной, причем единственной, развития всех фаз синаптического потенциала. При этом ацетилхолин очень быстро диффундирует в синаптической щели и сразу связывается с рецепторами на поверхности мышечной клетки. Впоследствии было установлено, что ацетилхолиновый рецептор представляет собой белок, состоящий из двух основных компонентов: структуры, связывающей ацетилхолин, и ионного канала. Когда рецептор узнает и связывает ацетилхолин, это вызывает открывание ионного канала.
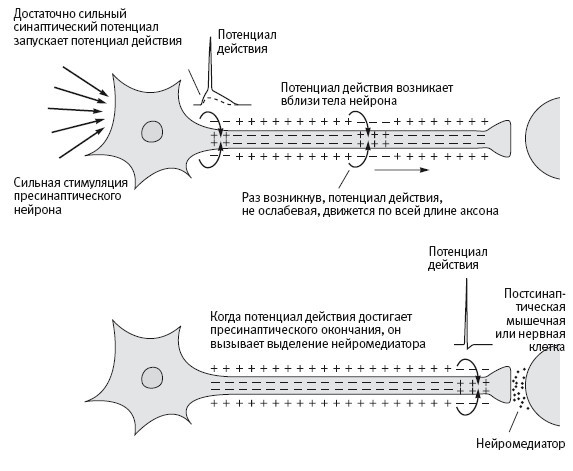
Затем Кац показал, что ионные каналы, зависимые от химического медиатора, отличаются от потенциал-зависимых натриевых и калиевых каналов двумя свойствами: они реагируют только на специфический химический медиатор и пропускают и натрий, и калий. Одновременное прохождение ионов натрия и калия по этим каналам меняет мембранный потенциал мышечной клетки от –70 милливольт почти до нуля. Кроме того, хотя синаптический потенциал и вызывается химическим путем, это происходит очень быстро, как и предсказывал Дейл. Если синаптический потенциал оказывается достаточно сильным, он вызывает потенциал действия, который приводит к сокращению мышечного волокна (рис. 6–2).
6–2. Распространения потенциала действия.
Результаты, полученные общими усилиями Ходжкина, Хаксли и Каца, доказали, что есть два принципиально разных типа ионных каналов. Потенциал-зависимые вызывают потенциалы действия, передающие информацию в пределах нейрона, в то время как медиатор-зависимые передают информацию между нейронами (или между нейронами и мышечными клетками), вызывая потенциалы действия в постсинаптических клетках. Таким образом, Кац выяснил, что медиатор-зависимые каналы, вызывая синаптический потенциал, как бы преобразуют химические сигналы, идущие от мотонейронов, в электрические, принимаемые мышечными клетками.
Наряду с болезнями, связанными с дефектами потенциал-зависимых каналов, существуют и болезни, связанные с дефектами медиатор-зависимых каналов. В частности, при миастении – серьезном аутоиммунном заболевании, встречающемся преимущественно у мужчин, – организм производит антитела, разрушающие ацетилхолиновые рецепторы мышечных клеток и тем самым ослабляя мышцы. Иногда мышцы ослабевают настолько, что пациент не может даже держать глаза открытыми.
Синаптическая передача в спинном и головном мозгу определенно сложнее, чем передача сигналов между мотонейронами и мышцами. Годы с 1925‑го по 1935‑й Экклс провел за исследованиями спинного мозга под непосредственным руководством Шеррингтона. В 1945 году он вернулся к этой работе и стал заниматься преимущественно ею и к 1951 году получил данные внутриклеточной регистрации мембранного потенциала мотонейронов. Экклс подтвердил вывод Шеррингтона, что мотонейроны получают как возбуждающие, так и тормозные сигналы и что эти сигналы передаются специфическими нейромедиаторами, воздействующими на специфические же рецепторы. Возбуждающие нейромедиаторы, выделяемые ведущими к мотонейрону пресинаптическими нейронами, повышают мембранный потенциал постсинаптической клетки с –70 до –55 милливольт – пороговой величины для запускания потенциала действия, в то время как тормозящие нейромедиаторы снижают мембранный потенциал от –70 до –75 милливольт, в результате чего запустить потенциал действия оказывается намного сложнее.
Как нам теперь известно, основным возбуждающим медиатором в головном мозгу служит глутаминовая аминокислота, а основным тормозящим – аминокислота ГАМК (гамма-аминомасляная кислота). Многие транквилизаторы (например, бензодиазепины, барбитураты, алкоголь и средства для наркоза) связываются с рецепторами ГАМК и оказывают успокаивающее действие на организм, усиливая торможение, обеспечиваемое этими рецепторами.
Тем самым Экклс подтвердил вывод Каца, что возбуждающая синаптическая передача имеет химическую природу, и доказал, что тормозная синаптическая передача тоже имеет химическую природу. Описывая впоследствии эти открытия, Экклс писал: “Карл Поппер убедил меня сформулировать свою гипотезу как можно конкретнее, чтобы она располагала к экспериментальной проверке на ложность. Случилось так, что ложность этой гипотезы мне удалось доказать самому”. Экклс отметил свои открытия тем, что отказался от электрической гипотезы, которую он так активно отстаивал, и стал искренним сторонником химической, столь же активно и увлеченно доказывая ее универсальность.
В это самое время, в октябре 1954 года, Пол Фэтт, один из талантливых соавторов Каца, подготовил превосходную обзорную работу о синаптической передаче. В этой работе Фэтт прозорливо отметил, что было преждевременно делать вывод, что синаптическая передача всегда имеет химическую природу. В заключение он написал: “Хотя все и указывает на то, что химическая передача происходит во всех соединениях <…> лучше всего знакомых физиологу, вполне возможно, что в некоторых других соединениях происходит электрическая передача” (курсив мой. – Э. К.).
Через три года справедливость предсказания Фэтта убедительно продемонстрировали Эдвин Фершпан и Дэвид Поттер, два постдока из лаборатории Каца, обнаружившие конкретный пример электрической передачи между двумя клетками в нервной системе рака. Таким образом, как это иногда и бывает с научными спорами, обе стороны оказались в чем‑то правы. Теперь мы знаем, что большинство синапсов, в том числе те, что исследовались во времена этого спора, имеют химическую природу. Но некоторые нейроны образуют с другими электрические синапсы. В таких синапсах между двумя клетками появляются небольшие мостики, позволяющие электрическому току проходить из одной клетки в другую – примерно так, как некогда предсказывал Гольджи.
Открытие двух форм синаптической передачи заставило меня задаться вопросами, к которым мне еще предстояло вернуться. Почему в мозге преобладают химические синапсы? Не разные ли роли играют в поведении химическая и электрическая передачи?
На последнем этапе своей выдающейся научной карьеры Кац оставил исследования синаптического потенциала клеток-мишеней и обратился к изучению выделения нейромедиатора передающей сигнал клеткой. Он стремился узнать, как электрическое явление в пресинаптическом окончании (потенциал действия) вызывает выделение химического медиатора. Он сделал в этой области два замечательных открытия. Во-первых, когда распространяющийся по аксону потенциал действия достигает пресинаптического окончания, это приводит к открыванию потенциал-зависимых каналов, впускающих в клетку ионы кальция. Приток ионов кальция внутрь пресинаптического окончания запускает серию молекулярных реакций, приводящих к выделению нейромедиатора. Таким образом, открываемые потенциалом действия потенциал-зависимые кальциевые каналы пресинаптической клетки запускают процесс преобразования электрического сигнала в химический, точно так же, как в принимающей сигнал клетке медиатор-зависимые каналы преобразуют химические сигналы обратно в электрические.
Во-вторых, Кац открыл, что медиаторы, такие как ацетилхолин, выделяются из окончания аксона не по одной молекуле, а отдельными небольшими порциями, примерно по пять тысяч. Кац назвал эти порции квантами и предположил, что каждая заключена в окруженный мембраной мешочек, который он обозначил как синаптический пузырек. Микрофотографии синапса, полученные в 1955 году Сэнфордом Пейли и Джорджем Паладе с помощью электронного микроскопа, подтвердили предположение Каца, показав, что пресинаптическое окончание набито пузырьками, в которых, как было доказано впоследствии, содержатся молекулы нейромедиатора (рис. 6–3).