Возможность последовательной разборки молекулы без повреждения остающихся частей – факт весьма нетривиальный. Все блоки молекулы связаны друг с другом, причем связи эти имеют направленный характер: при их разрыве один блок повреждается, а другой – нет. Можно представить систему блоков и связей между ними как множество точек, соединенных стрелками, причем стрелка будет указывать на тот блок, который повреждается при разрыве связи. Если бы эти стрелки образовали хотя бы одну кольцевую структуру (иными словами, если бы мы, двигаясь из какой-то точки по стрелкам, могли вернуться в ту же точку), то разобрать молекулу без повреждения остающихся частей было бы невозможно. Однако ни одной такой кольцевой структуры в молекуле 23S-pPHK не обнаружилось. Если бы направление связей было случайным, вероятность отсутствия кольцевых структур составляла бы менее одной миллиардной. Значит, это вряд ли результат случайности. По-видимому, структура связей между блоками молекулы отражает последовательность добавления этих блоков в ходе постепенной эволюции молекулы.
Получается, что исходной функциональной молекулой – "проторибосомой", с которой началась эволюция рибосомы, – был каталитический центр молекулы 23S-pPHK, ответственный за соединение аминокислот.
Могла ли такая "проторибосома", способная удерживать две молекулы тРНК и сближать в пространстве прикрепленные к ним аминокислоты, выполнять какую-то полезную функцию в РНК-организме? Эксперименты позволяют ответить на этот вопрос утвердительно. Методом искусственной эволюции были получены функциональные РНК (рибозимы), способные катализировать соединение аминокислот, прикрепленных к тРНК, в короткие белковые молекулы. Структура этих искусственно выведенных рибозимов очень близка к структуре той проторибосомы, которую "вычислили" канадские биохимики на основе изучения структуры 23S-pPHK.
По-видимому, проторибосома была просто устроенным рибозимом, катализирующим синтез небольших белковых молекул в РНК-организме. Специфичность синтеза поначалу была очень низкой (аминокислоты выбирались более или менее случайно). В дальнейшем к проторибосоме добавлялись новые блоки, причем добавлялись они таким образом, чтобы не нарушить структуру активного центра молекулы, а также всех тех блоков, которые присоединились ранее. Если очередная мутация приводила к нарушению уже сложившихся структур, она отсеивалась отбором.
Ученые детально реконструировали предполагаемый процесс постепенной эволюции 23S-pPHK. Первые восемь дополнительных блоков присоединились к проторибосоме таким образом, что образовали нечто вроде массивного "основания", благодаря которому структура проторибосомы стала гораздо более стабильной. Следующие 12 блоков еще более укрепили и расширили это "основание". Новые блоки образовали поверхность контакта с малой субъединицей, что позволило включить ее в состав рибосомы. В числе последних добавились блоки, образующие особые выросты на поверхности большой субъединицы. Функция этих выростов состоит в том, что они помогают рибосоме выбирать "правильную" тРНК, несущую нужную аминокислоту, а также выпускать из рибосомы "отработанные" тРНК. В итоге проторибосома оказалась окружена другими блоками со всех сторон за исключением канала, который был оставлен для выхода образующейся белковой молекулы.
Таким образом, 23S-pPHK, при всей ее кажущейся сложности, построена на основе довольно простого принципа. Ее блочная структура свидетельствует о том, что она могла довольно быстро развиться в ходе эволюции из проторибосомы под действием мутаций и отбора.
ДНК и РНК – хранители наследственной информации. ДНК, как и РНК, формируются из кирпичиков – нуклеотидов, только чуть-чуть других. Чтобы из РНК-кирпичика (рибонуклеотида) сделать ДНК-кирпичик (дезоксирибонуклеотид), достаточно одной простой реакции – отнять у рибозы один из атомов кислорода. Это придает молекуле стабильность, а заодно лишает ее способности совершать активные действия.
Молекулы ДНК (как и РНК) способны к самокопированию, правда, для этого нужны катализаторы – белки или рибозимы. Наследственная информация, хранящаяся в ДНК в виде последовательности нуклеотидов, может "переписываться" на РНК (так создаются матричные РНК, мРНК) и обратно. Точность копирования обеспечивается в значительной мере автоматически – благодаря особому свойству нуклеотидов, которое называют свойством комплементарности: против каждого нуклеотида исходной молекулы (матрицы) в синтезируемой копии (реплике) может встать только один, строго определенный нуклеотид из четырех возможных. Напротив гуанина (Г всегда становится цитозин (Ц), напротив урацила (У) или замещающего его в молекуле ДНК тимина (Т) – только аденин (А). Когда на этой реплике синтезируется новая реплика, она окажется точной копией исходной молекулы.
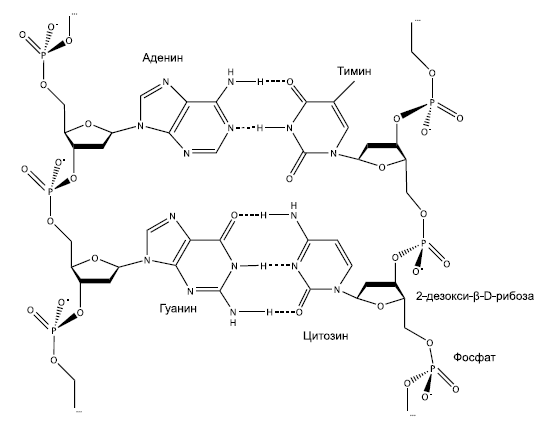
Фрагмент двойной цепи ДНК. По краям – "скелеты" одиночных цепей, составленные из остатков дезоксирибозы и фосфорной кислоты. В центре – две пары азотистых оснований, соединенных друг с другом по принципу комплементарности. Между аденином (А) и тимином (Т) образуются две водородные связи, между гуанином (Г) и цитозином (Ц) – три. Поэтому аденин может склеиться только с тимином, а гуанин – только с цитозином.
"Взаимовыгодное сотрудничество" РНК и белков (пептидов), вероятно, складывалось постепенно. Можно предположить, что изначально химические циклы с участием РНК и пептидов формировались порознь, возможно, в несколько разных условиях. Об этом свидетельствует химический состав этих веществ. В РНК много фосфора: остатки фосфорной кислоты вместе с остатками рибозы составляют "скелет" молекулы. Сера в состав РНК не входит. В белках, наоборот, нет фосфора, зато имеется сера, которая играет весьма важную роль в поддержании пространственной структуры белковой молекулы.
Поначалу синтез белков, осуществлявшийся РНК-организмами, скорее всего, не был строго специфичным: последовательности аминокислот из раза в раз воспроизводились не точно, а лишь приблизительно. Поскольку точность в данном случае резко повышала жизнеспособность организмов, естественный отбор способствовал выработке все более точных методов синтеза белка. Дело кончилось возникновением универсальной системы очень точного синтеза любого требуемого пептида. Это и был генетический код вкупе с рибосомами "современного" типа.
Генетический код – универсальный для всех живых существ способ, посредством которого первичная структура белковой молекулы (последовательность аминокислот) "кодируется" в молекуле ДНК (или РНК). Каждая аминокислота кодируется тремя нуклеотидами (кодоном, или триплетом). Нуклеотидов в ДНК всего 4, поэтому они могут образовывать 64 разных триплета. Аминокислот в белках всего 20, поэтому генетический код "избыточен": многие аминокислоты кодируются не одним, а несколькими взаимозаменимыми кодонами. Считывание генетической информации происходит в два этапа. Сначала информация "переписывается" с ДНК на РНК (транскрипция). Эту операцию осуществляет специальный фермент – ДНК-зависимая РНК-полимераза. Полученная в результате транскрипции молекула РНК, содержащая "инструкцию" по синтезу белка, называется матричной РНК (мРНК). Выполнение этой "инструкции", то есть синтез белка (трансляция), осуществляется рибосомами.
Вторым крупным усовершенствованием РНК-организмов было приобретение ДНК. Молекулы ДНК более устойчивы, чем РНК, и потому являются более надежными хранителями наследственной информации. Платой за стабильность стала неспособность молекул ДНК сворачиваться в сложные трехмерные структуры и выполнять какие-либо активные действия. Изначально ДНК, скорее всего, была чем-то вроде покоящейся фазы в жизненном цикле самовоспроизводящихся колоний РНК, и лишь много позднее она стала основным носителем наследственной информации.