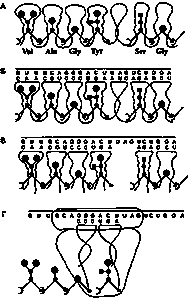
Рис. 1. Схемы, иллюстрирующие гипотезу перехода от примитивного синтеза предетерминированных пептидов к современному генетическому коду.
А. Образование белка (пептида), запрограммированного последовательностью петель-полостей единой молекулы РНК, связывающих активированные аминокислоты. Б. То же. Показана также “огибающая” РНК, нуклеотидная последовательность которой соответствует (комплементарна) ряду “торцов” петлевых элементов матрицы. В. Синтез пептидов на последовательности разделенных полостей-петель, набираемых из общего пула. Порядок петлевых элементов в наборе определяется связыванием их торцевых участков с соответствующими участками «огибающей» РНК, выполняющей таким образом роль информационной молекулы (мРНК). Соответственно, петлевые элементы в этой схеме являются предшественниками тРНК. Г. Эпизод современного синтеза белка на рибосоме: присоединение очередного звена к растущей белковой цепи. Вследствие изменения механизма узнавания адапторами соответствующих аминокислот петли тРНК перестали представлять собою «полости», хотя в них присутствуют необходимые для узнавания структурные элементы.
Схемы Рис. 1 предназначены для иллюстрации основных положений гипотезы и не отражают истинной конформации (укладки) матричных молекул РНК. Другие разъяснения приведены в тексте. Рисунок создан при участии В.В. Горбенко.
Переход к современному принципу кодирования аминокислотных последовательностей и к способу прочтения этого кода при синтезе белка в рибосоме иллюстрирует схема Рис. 1Б: связавшие аминокислоты петли-адапторы выстроены в том же порядке, что на Рис. 1А и Б, но не потому, что они в этом порядке связаны друг с другом в единой молекуле, а потому, что получили информацию о своей позиции благодаря взаимодействию с соответствующими участками огибающей РНК, принявшей на себя функции мРНК. Участки, узнаваемые в мРНК адапторными РНК, представляли собою кодоны, а их последовательность кодировала определенный пептид.
Поначалу новый способ кодирования белков развивался как вспомогательный вариант “под прикрытием” продолжавшего функционировать примитивного механизма. Главным эволюционным стимулом к переключению на новый способ кодирования белков явилась высокая рациональность этого способа, освободившего клетку от необходимости содержать и сохранять в поколениях значительное количество (по числу белков) РНК-матриц, размеры которых многократно превышали размеры соответствующих мРНК. После перехода к кодированию с участием мРНК открылась возможность увеличения как числа, так и размеров клеточных белков. Соответственно возросли их разнообразие и конформационная сложность, что позволило клеткам освоить новые пути метаболизма, усовершенствовать энергетику, кардинально повысить скорость и точность синтезов.
Первоначально характер связывания аминокислот соответствующими им адапторами сохранялся таким же, как при примитивном синтезе: полость, образованная петлей РНК, конформационно соответствовала аминокислоте и удерживала ее в связанном положении. Впоследствии механизм связывания был изменен. Оно стало осуществляться при посредничестве белковых ферментов аминоацил-тРНК-синтетаз, подключающих аминокислоты к 3'-концевому аденозину соответствующего адаптора (тРНК). При этом изменилось назначение петлевой области в молекулах тРНК. Ее структура сохранила специфичность, но преобразовалась соответственно новому назначению — служить объектом узнавания для специфической аминоацил-тРНК-синтетазы.
Предлагаемая модель позволяет удовлетворительно объяснить происхождение ряда характерных особенностей современного генетического кода, в частности, появление “бессмысленных кодонов”, обрывающих синтез пептида на рибосоме. При синтезе белка по единой РНК-матрице их роль выполняли петли, которые вообще не связывали аминокислоту (Рис. 1А). Такие петли разделяли два пептида, образованные на единой РНК-матрице. В пуле разделившихся петель-адапторов “пустой” петлевой элемент не закрепился за ненадобностью, но соответствующий ему кодон сохранился в мРНК (Рис. 1Б).
В схемах Рис. 1, иллюстрирующих гипотезу возникновения современного способа кодирования белков, использован нынешний трехнуклеотидный (трехбуквенный) код. Однако на раннем этапе перехода к современному способу кодирования код, вероятно, был иным. Логично предположить, что число букв в нем было не три, а не менее чем семь-девять. Благодаря этому энергия комплементарного кодон-антикодонного взаимодействия могла обеспечить стабильность комплекса на время, необходимое для образования пептидной связи.
Большой размер раннего кодона мог также в отсутствие специального механизма обеспечивать соблюдение рамки считывания. При трехбуквенном коде все возможные 64 триплета задействованы, т. е. за исключением трех стоп-кодонов они могут быть узнаны соответствующими тРНК. Поэтому смещение рамки считывания в мРНК, кодирующей определенный пептид, на одну или две буквы не прерывало бы синтеза, но изменило бы последовательность кодонов, т. е. привело бы к появлению “неправильного” пептида. При современном синтезе белка на рибосомах осуществляется контроль начала считывания со стартового кодона, определяющего N-концевую аминокислоту и одновременно обозначающего начало рамки считывания. Однако трудно рассчитывать на то, что контроль соблюдения рамки считывания уже осуществлялся в ранних версиях современного способа кодирования. Роль контролирующего фактора в соблюдении рамки считывания могли сыграть большие размеры кодона. При семибуквенном коде и четырех узнаваемых элементах (азотистых основаниях) число возможных вариантов кодонов около 16 000. Очевидно, что число функционировавших РНК-адапторов и, соответственно, “осмысленных” (соответствовавших определенным аминокислотам) кодонов было многократно ниже. Абсолютное большинство потенциальных кодонов не имело адапторов. Поэтому вопрос об использовании “неправильной” рамки считывания вообще не стоял: существовала единственная рамка, обеспеченная адапторами на всем протяжении. В ней осуществлялся синтез запрограммированного пептида.
Впоследствии, когда сформировался действующий поныне аппарат синтеза белков (Рис. 1В), включающий рибосому, которая наряду с другими функциями осуществляет узнавание стартового кодона в мРНК, контролирует последовательное подключение “заряженных” аминокислотами тРНК и стабилизирует кодон-антикодонное взаимодействие до момента формирования пептидной связи, размер кодона был редуцирован до необходимого минимума — триплета. Возможный ход эволюции генетического кода рассматривался ранее (Fitch and Upper, 1987).
3.3. Трансформация мира РНК в мир РНК-ДНК
Новый скачок в эволюции клетки связан с появлением ДНК и переходом к ней функций основного носителя генетической информации. Эволюционно этот переход был обусловлен меньшей склонностью ДНК к гидролизу (Бреслер, 1963) и, соответственно, более высокой прочностью полинуклеотидной цепи. Последнее обстоятельство позволяет ДНК формировать значительно более длинные молекулы, чем это возможно у РНК. В современной клетке только ДНК является автореплицирующейся молекулой и присутствует в форме двунитевой молекулы (биспирали). Все клеточные РНК синтезируются в форме однонитевых молекул путем комплементарного копирования “смысловой” нити двунитевой ДНК-матрицы.
Именно двунитевая структура ДНК повлекла формирование целой серии репарационных систем, способных узнавать и устранять дефекты в одной нити, используя информацию, которую несет комплементарная нить. Эти преобразования в структуре и метаболизме клетки, обеспечившие повышенную устойчивость ее генетического аппарата, были реализованы в хромосомах, содержащих двойную нить ДНК протяженностью в несколько миллионов нуклеотидных звеньев. Резко возросло и перестало быть лимитирующим фактором в эволюции общее количество генетического материала, которым способна управлять клетка (т. е. сохранять, реплицировать, эквивалентно распределять в потомстве, контролировать считывание информации). В современном мире на первом месте — скорость и контролируемость метаболического процесса. Поэтому в генах, помимо структурных участков, кодирующих белки или функциональные РНК (транспортные, рибосомные и др.), присутствуют регуляторные области, которые при участии белков активаторов и репрессоров контролируют транскрипцию.