Мутации наследуются так же, как исходные, неизмененные признаки. Например, в рассмотренном выше первом урожае ячменя может появиться несколько колосьев с остями, заметно выходящими за рамки изменчивости, показанные на рис. 7, вообще без ости. Они могут быть носителями мутации де Фриза, которая передастся потомству, а значит, все их потомки будут лишены ости.
Таким образом, мутация представляет собой изменение наследия и должна определяться неким изменением наследственного материала. На самом деле, большинство важных экспериментов по скрещиванию, объяснивших нам механизм наследственности, заключались в анализе потомства, полученного скрещиванием согласно разработанной схеме, носителей мутации (во многих случаях – нескольких мутаций) с немутантными особями или с носителями других мутаций. Кроме того, поскольку мутации наследуются, они являются подходящим материалом, с которым может работать естественный отбор, производя новые виды, как описал Дарвин, путем вымирания неприспособленных и выживания сильнейших. Нужно лишь заменить в теории Дарвина «незначительные случайные вариации» на «мутации» (так же, как квантовая теория заменяет «постоянный перенос энергии» «квантовым переходом»). Во всех прочих аспектах теория Дарвина нуждалась в незначительной корректировке, если я правильно понимаю точку зрения большинства биологов. Активно обсуждался вопрос, способствует ли естественному отбору (или даже доминирует ли над ним) явная склонность мутаций происходить в благоприятном направлении. У меня на данный момент нет своей точки зрения, однако следует отметить, что возможность «направленности» мутаций далее не учитывается. Более того, я не могу уделить внимание взаимодействию генов-«переключателей» и «полигенов», как бы ни были они важны для механизма естественного отбора и эволюции.
Локализация, доминантность и рецессивность
Теперь следует рассмотреть фундаментальные факты и понятия, касающиеся мутаций, вновь в немного категоричной форме, не затрагивая напрямую подтверждающие их экспериментальные доказательства.
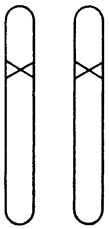
Рис. 8. Гетерозиготный мутант. Крестиком показан ген с мутацией
Разумно предположить, что некую наблюдаемую мутацию вызывает изменение определенного участка одной из хромосом. Так и есть. Важно отметить, что мы точно знаем: это изменение затрагивает лишь одну хромосому, но не соответствующий «локус» на гомологичной хромосоме. Это схематично показано на рис. 8; мутировавший локус обозначен крестиком. Факт, что мутация происходит только в одной хромосоме, можно доказать, скрестив мутантную особь («мутанта») с нормальной. Половина потомства будет обладать мутантным признаком, а другая половина – нормальным. Этого следует ожидать вследствие расхождения двух хромосом у мутанта во время мейоза, как схематично показано на рис. 9. Это «фамильное дерево», на котором каждая особь (из трех последовательных поколений) представлена парой рассматриваемых хромосом. Имейте в виду, что если у мутанта затронуты обе хромосомы, все его потомки получат одинаковое (смешанное) наследие, отличное от родительского.
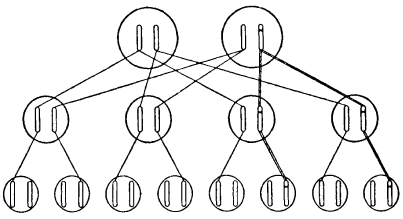
Рис. 9. Наследование мутации. Поперечные прямые линии показывают передачу хромосомы, двойные линии – передачу мутантной хромосомы. Неучтенные хромосомы третьего поколения пришли от партнеров второго поколения, не включенных в схему. Предполагается, что это не родственники, лишенные мутаций
Однако эксперименты в данной области не столь просты, как может показаться. Их затрудняет второй важный фактор, а именно то, что мутации очень часто латентны. Что это означает?
У мутанта две «копии кода» уже не идентичны, а представляют собой два различных «прочтения», или «версии», по крайней мере в одном конкретном месте. Следует отметить, что, как бы заманчиво это ни выглядело, нельзя считать исходную версию «ортодоксальной», а мутантную – «еретической». В принципе мы должны полагать, что они имеют равное право на существование, поскольку нормальные признаки также возникли благодаря мутациям.
Рис. 10. Гомозиготный мутант, представленный одной четвертью потомства, появившегося путем самооплодотворения гетерозиготного мутанта (рис. 8) либо скрещивания двух гетерозиготных мутантов
В действительности «структура» особи, как правило, определяется той или иной версией, которая может быть нормальной либо мутантной. Определяющую версию называют доминантной, другую – рецессивной. Иными словами, мутация является доминантной или рецессивной в зависимости от того, меняет ли она непосредственно структуру или нет.
Рецессивные мутации встречаются намного чаще доминантных и крайне важны, хотя поначалу никак не проявляют себя. Чтобы повлиять на структуру, они должны присутствовать на обеих хромосомах (см. рис. 10). Подобные особи появляются, когда друг с другом скрещиваются два одинаковых рецессивных мутанта или когда мутант скрещивается сам с собой. Такое бывает у гермафродитных растений и даже случается спонтанно. В данном случае около одной четверти потомков будут носителями двух мутантных хромосом, а значит, у них проявится мутантный признак.
Немного технических терминов
Полагаю, для ясности следует объяснить несколько технических терминов. То, что я назвал версией кода – исходной или мутантной, – называют аллелью. Если версии различаются, как на рис. 8, особь называют гетерозиготной по данному локусу. Если версии одинаковы, как у немутантной особи или в случае, показанном на рис. 10, такие организмы называют гомозиготными. Так, рецессивная аллель влияет на структуру только в гомозиготном состоянии, в то время как доминантная аллель дает одну и ту же структуру и в гомозиготном, и в гетерозиготном состоянии.
Цвет часто доминирует над бесцветностью (или белизной). Например, горох обладает белыми цветками, только когда содержит в обеих соответствующих хромосомах «рецессивную аллель, отвечающую за белизну», то есть когда он «гомозиготен по белизне». Этот признак будет наследоваться, и все потомки данного растения станут цвести белым цветом. Однако присутствие одной «красной аллели» (в то время как другая остается белой – у «гетерозиготы») – равно как и две «красных аллели» (у «гомозиготы») – заставит горох цвести красными цветками. Различие последних двух случаев проявится только в потомстве, когда красные гетерозиготы дадут несколько белых потомков, а красные гомозиготы сохранят признак.
Факт, что две особи могут обладать совершенно одинаковой внешностью, но разной наследственностью, является столь важным, что следует точно разграничить эти понятия. Генетики говорят, что у таких особей одинаковый фенотип, однако разный генотип. Таким образом, вышеизложенное можно суммировать кратким, но в высшей степени техническим утверждением: рецессивная аллель влияет на фенотип лишь в том случае, когда генотип гомозиготен.
Мы будем использовать эти технические термины, однако при необходимости будем напоминать читателю их значение.
Опасное влияние близкородственного скрещивания
В гетерозиготном состоянии рецессивные мутации – неподходящий материал для естественного отбора. Если они вредны, как часто и бывает, то все равно сохраняются в силу своей латентности. Поэтому значительное количество неблагоприятных мутаций может накапливаться, не причиняя непосредственного вреда. Однако они передаются половине потомков – и это имеет большое значение для человека, домашнего скота, птицы и любого другого вида, физическое состояние которого нас заботит. На рис. 9 предполагается, что особь мужского пола, например я сам, несет рецессивную вредную мутацию в гетерозиготном состоянии, то есть она не проявляется. Предположим, у моей жены нет такой мутации. Тогда половина наших детей (второе поколение) окажется носителями мутации – снова гетерозиготной. Если все они опять выберут себе партнеров без мутаций (не показаны на схеме для ясности), около четверти наших внуков будут обладать гетерозиготной мутацией.