Гаплоидные особи
Еще один момент требует уточнения. Для нас он не слишком важен, однако представляет значительный интерес, поскольку демонстрирует, что практически полный код «структуры» содержится в каждом одинарном наборе хромосом.
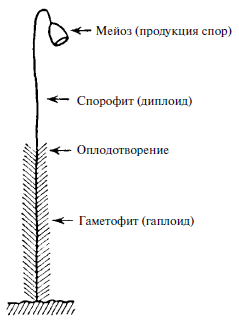
Рис. 5. Чередование поколений
В некоторых случаях непосредственно после мейоза не происходит оплодотворения, и гаплоидная клетка («гамета») претерпевает многочисленные митотические деления, приводящие к формированию полностью гаплоидной особи. Например, самец пчелы – трутень – развивается партеногенетически, то есть из неоплодотворенных, а значит, гаплоидных яйцеклеток матки. У трутня нет отца! Все клетки его тела гаплоидны. Можно назвать его огромным сперматозоидом – и действительно, как известно, это его единственная функция. Однако данная точка зрения нелепа. Такой случай не уникален. Существуют семейства растений, у которых гаплоидная гамета, образующаяся путем мейоза и называемая спорой, падает на землю и, подобно семени, развивается в истинно гаплоидное растение, по размерам сравнимое с диплоидным. На рис. 5 приведено схематическое изображение мха, распространенного в наших лесах. Облиственная нижняя часть представляет собой гаплоидное растение, называемое гаметофитом, потому что на его верхнем конце развиваются половые органы и гаметы, которые путем взаимного оплодотворения дают диплоидное растение – голый стебель со споровой коробочкой наверху. Когда коробочка раскрывается, споры попадают на землю, из них развивается облиственный стебель и т. д. Эту последовательность событий называют чередованием поколений. Можно взглянуть подобным образом на человека и животных. Однако «гаметофит» у них, как правило, представляет собой короткоживущее одноклеточное поколение, сперматозоид или яйцеклетку. Наше тело соответствует спорофиту. Наши «споры» – запасенные клетки, из них посредством мейоза образуется одноклеточное поколение.
Значимость редукционного деления
Важным, действительно судьбоносным событием в процессе размножения особи является не оплодотворение, а мейоз. Один набор хромосом достается от отца, другой – от матери. Ни случай, ни рок не может повлиять на это. Каждый человек[15] обязан половиной своего наследственного материала отцу и половиной – матери. То, что обычно одна из них доминирует, объясняется другими причинами, до которых мы еще дойдем. Разумеется, самый простой пример такого доминирования – пол.
Но если проследить вашу наследственность до бабушки и дедушки, ситуация изменится. Возьмем набор хромосом моего отца, точнее, одну их пару, например № 5. Это верная копия № 5, которую отец получил от своего отца, или № 5, какую отец получил от своей матери. Вопрос был решен с вероятностью 50:50 в ходе мейоза, случившегося в организме моего отца в ноябре 1886 года и произведшего сперматозоид, который несколько дней спустя принял участие в зачатии меня. Аналогичную же историю можно рассказать про хромосомы № 1, 2, 3… 24 из отцовского набора и, mutatis mutandis[16], из материнского набора. Более того, все 48 случаев независимы друг от друга. Даже если бы я знал, что отцовская хромосома № 5 попала ко мне от моего деда Йозефа Шредингера, хромосома № 7 имеет равные шансы также принадлежать ему – или его жене Марии, в девичестве Богнер.
Кроссинговер. Местоположение признаков
Однако чистая случайность получила еще больше возможностей смешать в потомке наследие дедов и бабок, чем может показаться на основании вышеизложенного. Ранее мы полагали, будто конкретная хромосома целиком досталась нам от деда или от бабки, иными словами, что отдельные хромосомы передаются неделимыми. В действительности это не так, по крайней мере, не всегда. Прежде чем разойтись в ходе редукционного деления, например, происходящего в организме отца, любые две «гомологичные» хромосомы вступают в близкий контакт друг с другом и обмениваются целыми участками, как показано на рис. 6. Благодаря этому процессу, называемому кроссинговер, два признака, расположенные в соответствующих частях данной хромосомы, окажутся разделены у внука, который получит одно из них от деда, а другое от бабки. Процесс кроссинговера снабдил нас важной информацией о расположении признаков на хромосомах. Для полноценного рассмотрения следовало бы очертить концепции, которые будут представлены в следующей главе (такие как гетерозиготность, доминантность и т. д.), однако это выведет нас за рамки данной книги, а потому позвольте сразу перейти к сути.
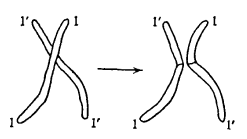
Рис. 6. Кроссинговер. Слева: контакт между двумя гомологичными хромосомами. Справа: гомологичные хромосомы после обмена и расхождения
Если бы не кроссинговер, два признака, за которые отвечает одна и та же хромосома, всегда передавались бы вместе, ни один потомок не смог бы получить один из них, не унаследовав при этом другой. Однако два признака, находящиеся на различных хромосомах, будут разделены с вероятностью 50:50 – или со 100 % вероятностью, если расположены на гомологичных хромосомах одного предка, которые не могут передаться вместе.
Кроссинговер нарушает эти правила. Таким образом, вероятность данного события можно оценить, тщательно отслеживая процентный состав потомства в длительных экспериментах по скрещиванию. Анализируя статистику, следует принять рабочую гипотезу, что «связь» между двумя признаками, расположенными на одной хромосоме, нарушается тем реже, чем ближе они находятся друг к другу. В таком случае ниже вероятность, что точка обмена окажется между ними, в то время как признаки, расположенные на противоположных концах хромосом, разделяются при каждом кроссинговере. То же самое верно для рекомбинации признаков, расположенных на гомологичных хромосомах одного предка. Таким образом, на основании «статистики связи» можно получить некую «карту признаков» каждой хромосомы.
Эти ожидания полностью подтвердились. В проведенных экспериментах, в основном – но не только – на дрозофиле, изученные признаки действительно разделяются на группы, не сцепленные друг с другом, число которых равняется числу отдельных хромосом (четыре у дрозофилы). В каждой группе можно составить линейную карту признаков, количественно соответствующую степени сцепления между любыми двумя признаками в данной группе. Поэтому нет сомнений, что они действительно расположены на хромосоме, причем линейно, что предполагает ее палочковидная форма.
Разумеется, изображенный здесь схематический механизм наследственности выглядит поверхностным и бесцветным, даже слегка наивным. Ведь мы не сказали, что именно называем «признаком». Кажется неправильным и невозможным расчленять на отдельные «признаки» структуру организма, который по существу является единым, «целым». В любом конкретном случае мы утверждаем, что пара предков различалась по явно заданным характеристикам (например, у одного были голубые глаза, а у другого – карие), и потомок в данном смысле похож на одного из них. То, что мы обнаруживаем в хромосоме, и есть причина этого различия. (Технический термин для этого – «локус», или, если имеется в виду лежащая в его основе гипотетическая структура, «ген».) На мой взгляд, различие в признаках само по себе является фундаментальной концепцией, а не признаком, вопреки очевидному лингвистическому и логическому противоречию данного утверждения. Различия в признаках дискретны, как мы поймем из следующей главы, когда будем обсуждать мутации, и я надеюсь, что приведенная здесь сухая схема обретет жизнь и заиграет красками.
Максимальный размер гена
Мы только что ввели термин «ген» – гипотетический материальный носитель определенной наследственной характеристики. Теперь следует подчеркнуть два момента, в высшей степени значимые для нашего исследования. Во-первых, размер – а точнее, максимальный размер – такого носителя; иными словами, до сколь малого объема мы сможем проследить местоположение признака? Во-вторых, постоянство гена, которое следует из устойчивости наследственной структуры.