Образование условного рефлекса кормления объясняет также, почему маленькие птицы выкармливают кукушонка, убивающего их детей. Это не «слепота инстинкта». Если подсадить кукушонка к не кормившей его птице, она улетит от него прочь.
Условные пищевые рефлексы подбегания к кормушке изучали в опытах с чижами и дубоносами. Такие рефлексы вырабатывались у них за 40–50 сочетаний, а у щеглов — за 70–80 сочетаний. После 2–3-месячного перерыва в работе для восстановления рефлекса требовалось около 20 сочетаний. Нетрудно было выработать дифференцировку по яркости светового сигнала. Однако на звуковые сигналы устойчивого условного рефлекса выработать не удавалось.
С канарейками, щеглами, синицами, снегирями и клестами проводили опыты в лабиринте с пищевым подкреплением. В этих опытах, например, снегирь при первой пробе искал выход 50 мин и прошел за это время 814 м, а после 40 упражнений находил выход за 1–2 мин. Ориентирами для птиц в лабиринте служили главным образом зрительные сигналы. Изменение условий освещения, приводящее к иному расположению света и теней, ослабляло вплоть до исчезновения всю выработку реакции выхода из лабиринта. Слабое освещение затрудняло поиски правильного пути, а темнота делала их безрезультатными. Вместе с тем смазывание стенок пахучими веществами, удлинение или укорочение коридоров не оказывало заметного влияния на поведение птиц в лабиринте. Следовательно, обоняние, осязание и мышечное чувство не имели существенного значения в поисках правильного пути. Рассказы о «говорящих» птицах часто грешат антропоморфизмом, т.е. птиц ошибочно наделяют человеческим сознанием. Так, попугаям (Psittaci), которые точно, вплоть до интонаций, могут имитировать голос человека, приписывают способность понимать смысл произносимых ими слов. Но тогда следовало бы ожидать, попугаи могут вести между собой и с людьми осмысленный разговор. Кроме того, пришлось бы допустить, что, например, пересмешники (Miminae), прекрасно воспроизводящие лай собаки, мычание коровы, ржание лошади, хрюканье свиньи, кукарекание петуха и т.д., понимают «языки» этих животных. Беспочвенность подобных предположений очевидна. Подражание птиц человеческому голосу не имеет ничего общего со словесным выражением мыслей у человека.
Хищные птицы. Орел, парящий под облаками, охватывает взором громадные просторы лесов и лугов, степей и гор и, заметив жертву, камнем падает на нее.
Мелкие соколы, охотящиеся за грызунами, например кобчик, могут с большой высоты заметить полевую мышь и безошибочно броситься на нее, увидеть одиночную стрекозу на расстоянии <...>00[7] м. Более крупных хищных птиц, таких, как кречет и сапсан, издавна использовали для охоты за дикими гусями, лебедями, цаплями.
Соколиная охота была широко распространена в Древней Руси, где искусство обучения боевых птиц стояло очень высоко. Сначала у сокола затормаживали натуральные, в основном зрительные, рефлексы, надевая на голову клобучок. Потом, удерживая сокола на привязи, у него вырабатывали ряд искусственных условных рефлексов схватывания добычи по сигналам посвистывания, прищелкивания и покрикивания. Постепенно круг усвоенных навыков расширялся, и обучение переносилось в поле. Хорошо подготовленный сокол летит на большой высоте за хозяином и бьет сверху вспугнутую дичь. В Средней Азии с помощью беркутов охотятся за лисицами, зайцами и даже волками. На Кавказе с помощью прирученных ястребов-перепелятников добывают перепелов на осеннем пролете.
Хищные птицы, даже питающиеся падалью, например коршуны или грифы-стервятники, в поисках пищи руководствуются в основном зрительными, отчасти слуховыми, но не обонятельными натуральными условными сигналами. Об этом говорят следующие опыты наблюдения.
В поле выставили чучело лани из хорошо просушенной кожи, набитое травой, и на него немедленно набросились грифы. В то же время труп свиньи, прикрытый сверху тростником, лежал несколько дней, стал разлагаться, но пролетавшие над ним грифы ничего не замечали. Один орнитолог приманивал рябчиков на пищик, имитирующий их голоса. На него налетел ястреб-перепелятник и ударил в голову, пытаясь схватить мнимого рябчика. Здесь хищник явно ориентировался только на слух.
Водоплавающие птицы. Домашние гуси и утки отличаются от диких сородичей не только особенностями телосложения, но и совершенно иным поведением. У них образовались новые инстинкты, искусственно созданные человеком. В то же время у диких видов сформировались другие натуральные реакции, определяемые условиями их жизни.
На примере поведения водоплавающих птиц наглядно проявляется значение для условно-рефлекторной деятельности принципа «биологической адекватности» раздражителей. Например, очень сильные, но не встречающиеся в естественных условиях звуки звонка почти не вызывали у дикого гуся двигательных реакций и изменений частоты сокращений сердца. Вместе с тем треск ломаемых щепок, напоминающий хруст, который производят, пробираясь через заросли (возможная опасность), заставлял селезня прятаться, замедляя ритм сердечных сокращений, а звуки плеска воды (приближение преследователя) или свист (бросающийся сверху пернатый хищник) приводили к бурной реакции бегства и резко учащали сердцебиение. Однако даже не действующий сам по себе звонок можно легко было превратить в сильно действующий сигнал, если каждое его 2-секундное применение сопровождать сильным безусловным раздражителем, например вдыханием паров аммиака. При этом после 3–5 сочетаний на звонок образуется условный оборонительный рефлекс с резким учащением частоты сердечных сокращений, который оказывается довольно прочным.
Натуральным сигналом половой реакции селезня является своеобразный крик самки. Притягательную силу этого рефлекса используют на охоте, когда диких самцов приманивают «подсадными» домашними утками или имитируют крик самки звуками манка. На основе таких натуральных рефлексов могут образовываться разнообразные двигательные и дыхательные условные рефлексы.
У диких и домашних (пекинских) уток способность к их выработке оказалась несколько различной. Например, условный рефлекс на метроном с частотой 200 ударов/мин образовался у дикой утки на 7-м сочетании, а у домашней — на 10-м. Дифференцировка метронома с частотой 90 ударов/мин была достигнута у дикой утки на 6-м его применении, а у домашней — лишь на 38-м. Следовательно, высшая нервная деятельность дикой утки может обеспечить более быстрые, чем у ее домашних сородичей, приспособительные реакции на меняющиеся условия жизненной обстановки.
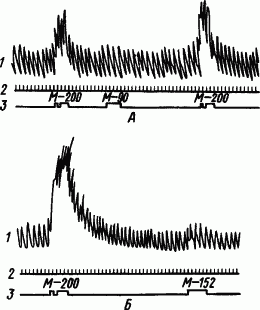
Однако острота слухового анализа даже у дикой утки оказалась сравнительно небольшой — выработать полную дифференцировку звуков метрономов 200 и 152 ударов/мин она была не в состоянии (рис. 87). Переделывая охотничьи инстинкты некоторых водоплавающих птиц, их используют для рыбной ловли.
Так, рыбаки южных морей обучают бакланов ловить рыбу и приносить ее хозяину. Для этого баклану надевают шею кольцо, препятствующее проглатыванию пойманной рыбы. Кольцо снимают лишь тогда, когда он принесет рыбу в лодку. Вначале баклана держат на привязи. Когда вырабатывается досрочно прочное торможение рефлекса проглатывания добычи и укрепляется положительный пищевой рефлекс на хозяина, то его выпускают в свободный полет. Вся охота превращается для баклана в цепь сигнальных пищедобывательных актов, за которыми следует пищевое подкрепление, когда, закончив ловлю, хозяин снимет с его шеи кольцо и покормит.
Рис. 87. Определение предела дифференцирования частоты стука метронома дикой уткой. А — полная дифференцировка; Б — неполная дифференцировка (по В.Г. Осиповой):
1 — дыхание, 2 — отметка времени, 3 — отметка сигналов