Активная природа отдельных разновидностей условного торможения обнаруживается в основных свойствах их проявлений. Например, у собаки были выработаны два условных слюнных рефлекса: на стук метронома и звук звонка. Если угашать рефлекс на стук метронома, а затем испытать рефлекс на звонок, то последний окажется уменьшенным. Угасательное торможение, возникшее <...пропущено слово...> первом рефлексе, распространилось на второй. Но ведь второй рефлекс «не работал» и в нем не может быть утомления. Следовательно, угасательное торможение не является результатом истощения нейронов.
Также не может быть следствием истощения запаздывательное торможение, так как по мере его выработки величина рефлекса растет, т.е. усиливается развиваемое клеткой условное возбуждение. Несовместимы с утомлением и свойства других разновидностей внутреннего торможения.
Но если внутреннее торможение является активным процессом, тяжелой работой, то как с этим примирить его восстановительное действие (например, в случае сна)? На первый взгляд, здесь явное противоречие. Однако противоречие исчезает, если вспомнить, что условное торможение вырабатывается. Можно предположить, что, вырабатывая торможение, нервная система напряженно работает, но когда торможение вырабатывается, она оказывается освобожденной от большей части своей деятельности и отдыхает.
Естественно, что отношения работы и отдыха при разных условиях развития торможения могут быть очень различными. Следует также иметь в виду, что при любом виде торможения лишь часть нервных клеток снижает свою импульсную активность, а другая часть ее увеличивает. Торможение нервного центра — не деятельность всех составляющих его нейронов, а активная деятельность, организованная особым образом из возбужденных и заторможенных клеток.
Локализация условного торможения. В каких анатомических отделах нервной системы может возникать и развиваться условное торможение? Ясный ответ на этот вопрос дает следующий факт. При повторении индифферентного раздражения у собаки происходит угашение ориентировочного рефлекса на этот раздражитель. Так как это торможение вырабатывается, следовательно, оно является внутренним. Если же проделать такой опыт с собакой, у которой целиком удалены большие полушария мозга, то угашения ориентировочного рефлекса не происходит. Без высших отделов мозга развитие угасательного, как и других форм приобретаемого торможения, становится невозможным. Поэтому безусловные рефлексы и протекают у бесполушарных животных так стереотипно. Осуществляющие их низшие отделы головного и спинного мозга сами не способны к выработке условных форм торможения, хотя и обнаруживают более простые свойства «привыкания». Однако они участвуют в его формировании путем воздействия на функциональное состояние коры главным образом через тормозящую таламокортикальную систему.
Выработка условного торможения так же, как и выработка временных связей условного рефлекса, происходит в высших отделах нервной системы; у высших позвоночных — в коре и подкорковых структурах больших полушарий мозга.
Но высшие отделы мозга, в частности кора больших полушарий, имеют сложную структуру. Условный рефлекс образуется благодаря временной связи между анализаторами сигнального раздражения и механизмом подкрепляющей реакции; в каком же звене нейронных сетей, образующих пути осуществления условного рефлекса, вырабатывается условное торможение?
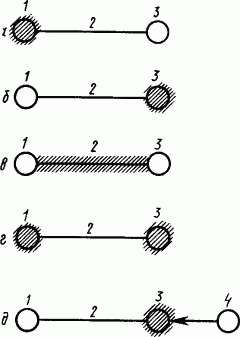
Результаты специальных исследований по изучению локализации условного торможения в корковых структурах мозга приводили ученых к разным взглядам (схематически представлены на рис. 31).
Рис. 31. Локализация внутреннего торможения (заштриховано) по представлениям различных авторов (по Л.Г. Воронину):
а — по Б.В. Бабкину, б — по И.Я. Перельцвейгу, в — по Э.А. Асратяну, г — по П.С. Купалову, д — по П.К. Анохину; 1 — корковая проекция условного раздражителя, 2 — временная связь, 3 — корковое представительство безусловного рефлекса, 4 — биологически отрицательная реакция
Первые исследования процесса угашения условного рефлекса, показавшие, что сигнальный стимул при определенных условиях перестает быть действующим, дали основание предполагать, что условное торможение развивается в зоне корковой проекции условного раздражителя (рис. 31, а). Дальнейшие опыты со слюноотделительными рефлексами на подкрепление вливанием в рот раствора кислоты привели к заключению о задержке сигналов в «кислотном центре» и выводу, что условное торможение развивается в зоне корковой проекции безусловного раздражителя (рис. 31, б). Так как на один и тот же сигнал можно было выработать два условных рефлекса: положительный и отрицательный, то, следовательно, зона проекции условного раздражителя не заторможена. В то же время на примере цепного двигательного рефлекса было показано, что зона проекции движении также остается назаторможенной. Это свидетельствовало в пользу мнения, что условное торможение развивается между зонами корковой проекции условного и безусловного раздражителей в структурах замыкания временной связи (рис. 31, в).
Исследование взаимодействия условных слюнных рефлексов при их выработке на базе безусловных, вызываемых раздражением выведенных наружу лоскутков слизистой языка, привело к заключению, что условное торможение развивается в зоне корковой проекции как условного, так и безусловного раздражителей (рис. 31, г). Наконец, исходя из того, что неподкрепление условного рефлекса вызывает «биологически отрицательную реакцию», которая может затормозить другую, одновременно протекающую деятельность, предположили, что условное торможение является внешним, которое «биологически отрицательная реакция» оказывает на зону корковой проекции безусловного раздражителя (рис. 31, д).
Систематические исследования функционального состояния всех звеньев структуры условного рефлекса при его угашении привели к заключению, что ни в одном из них не возникал очаг торможения, который мог блокировать условное возбуждение, и оно в зоне проекции сигнального раздражителя становилось источником внутреннего торможения. Этому соответствует тот факт, что нейронные реакции коры мозга кошки при выработке дифференцировочного торможения оказались различными в зонах условного и безусловного раздражителей.
Разногласия по вопросу о том, в каком звене механизма условного рефлекса развивается условное торможение, крайне затрудняют суждение о его нейронной структуре. Однако можно с большой долей вероятности принять, что оно протекает в тех же нервных клетках, которые в свое время участвовали в образовании временной связи. Об этом свидетельствует, к примеру, такой факт, что при угасательном торможении корковый нейрон замедляет свой разряд с тем же латентным периодом, с которым он учащал разряд при положительном рефлексе. В опытах с клеточным аналогом временной связи выработка дифференцировочного торможения изменяла лишь ту часть импульсной реакции нейрона, которая составляла его условный ответ. В процессе развития внутреннего торможения существенную роль играет взаимодействие корковых элементов с подкорковыми, в частности со структурами неспецифической системы таламуса и ретикулярной формации среднего мозга.
Электрофизиологические показатели условного торможения. Когда условный рефлекс угашается, он просто прекращается. В этом является конечный результат деятельности торможения, но не сама деятельность. Некоторые стороны этой деятельности, скрытой от внешнего наблюдения, находят свое отражение в динамике ритмов основной электрической активности. Если у кошки с вживленными электродами вырабатывать на метроном 120 ударов/мин дифференцировку условного пищевого рефлекса подбегания к кормушке, образованного на метроном 60 ударов/мин, то произойдут характерные изменения в картине электрических потенциалов. При этом положительный условный раздражитель будет продолжать подавлять колебания основного ритма (6–8 колебаний/с) электрической активности и усиливать более быстрые колебания (30–40 колебаний/с). В то же время дифференцировочный раздражитель вскоре перестанет вызывать такие изменения основной электрической активности.