Соотношение нейронных реакций, отражающих центральные процессы условного возбуждения, оказывается различным в областях коры, осуществляющих разные функции.
Таблица 1. Распределение возбуждающихся и тормозящихся нейронов в разных корковых областях мозга обезьяны при действии условного светового раздражителя (по Г. Джасперу и др.)

Как видно из табл. 1, в зонах проекции условного раздражителя и представительства двигательной реакции возбуждающихся нейронов зафиксировано примерно втрое больше, чем тормозящихся, а в зонах коры, выполняющих ассоциативные и интегративные функции (теменных и лобных), преобладали тормозящиеся нейроны, которых было почти вдвое больше, чем возбуждающихся. Видимо, для осуществления интегративных функций, более сложных, чем начальная обработка информации, в первичных проекционных зонах возрастает роль торможения, формирующая тонкую мозаику нейронной активности. В опытах на кошках также оказалось, что в областях, не являющихся зонами проекции условного и безусловного раздражителей, преобладают тормозящиеся нейроны. Однако в опытах на кроликах более половины нейронов и в зоне представительства условного слухового раздражителя отвечали на сигнал торможением. Доля нейронов, реагирующих торможением на сигнал, увеличивалась и в первичной соматосенсорной зоне мозга крысы при выработке мигательного рефлекса.
Роль тормозящихся нейронов в процессе образования условного рефлекса может заключаться в ограничении числа действующих синаптических связей, выделении специфических рецептивных полей и, тем самым, выделении необходимой информации из исходного шума. Вместе с тем анализ межнейронных связей в слуховой коре кошки показал, что при выработке условного рефлекса на звук усиливались возбудительные связи.
Характер условных реакций нейронов во многом зависит от организации их рецептивных полей. Так, нейроны, получающие сигналы от ограниченных полей, например кожной рецепции, оказываются наиболее стабильно реагирующими и мало меняют свои ответы на афферентное раздражение, когда он становится условным. Нейроны, получающие сигналы от распространенных полей рецепции, реагируют более вариабельно в связи с динамикой формирования условного рефлекса, что указывает на их более близкое отношение к механизму образования временных связей.
Так, в двигательной области коры кролика «обучающихся» нейронов было 78 % среди бимодальных, т.е. отвечающих на два рода раздражений, и только 34 % среди мономодальных, т.е. отвечающих лишь на раздражения одного рода. В коре мозга кошки 90 % полимодальных нейронов являются «обучающимися».
Способность нейронов включаться в условные реакции в значительной мере зависит от уровня возбудимости, о которой можно судить по частоте их фоновой импульсации. Именно нейроны высокой возбудимости, на которые сходятся сигналы от многих полей рецепции, играют важную роль в механизме образования временных связей. Вероятно поэтому нейронов, проявляющих условные реакции, в первичных проекционных полях оказалось меньше, чем во вторичных, где сопоставляется информация, приходящая по многим путям.
Так, в первичном зрительном поле 17 «обучающихся» нейронов было лишь 18 %, в то время как во вторичном зрительном поле 18 их оказалось 68 %. Вместе с тем «обучающиеся» клетки в большем или меньшем количестве можно найти в любой структуре мозга.
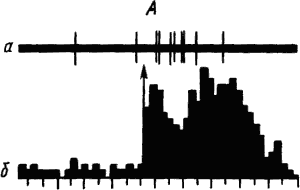
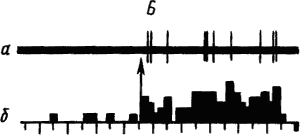
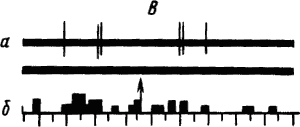
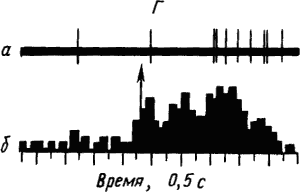
Активность «обучающихся» нейронов изменяется в соответствии со степенью выраженности внешних проявлений условного рефлекса и зависит от факторов, влияющих на эти проявления. Например, насыщение животного пищей ведет к уменьшению проявлений его пищевых условных рефлексов и резкому снижению условных реакций корковых нейронов. При инструментальных условных рефлексах такая зависимость наглядно показана для нейронов лобных отделов коры мозга обезьяны (рис. 19).
Рис. 19. Участие нейрона лобной коры обезьяны в реализации инструментального условного пищевого рефлекса в зависимости от потребности в пище. А — исходное состояние; Б — после еды; В — сытое животное не реагирует на сигнал; Г — другой вид пищевого подкрепления восстанавливает рефлекс:
а — нейронограмма, б — перистимульная гистрограмма, по оси координат — количество импульсов, стрелкой отмечен пусковой сигнал рефлекса
Странность, на первый взгляд, условных ответов корковых нейронов на действие сигнального раздражителя состоит в том, что их латентные периоды оказывались более длительными, чем при применении этого раздражителя как индифферентного (до сочетаний с безусловным). Так, некоторые корковые нейроны кошки при выработке оборонительного условного рефлекса на звуковой сигнал могли увеличивать латентный период своего импульсного ответа более чем втрое, от 30 до 100 мс. Для объяснения этого феномена, вероятно, следует исходить из того факта, что включение нейронов в механизм временной связи меняет их положение в нейронных сетях проведения возбуждения и состояние синаптической проводимости. Гистограммный анализ показал, что удлинение латентного периода получено в среднем из сложения удлиненных латентных периодов реакций одних нейронов и укороченных латентных периодов реакций других нейронов. Поэтому можно думать, что при формировании временных связей происходит наряду с усложнением путей передачи сигналов образование нейронных цепочек облегченного их проведения.
На этапе генерализации вырабатываемого условного рефлекса начинают вовлекаться нейроны обширных областей коры и подкорковых структур, гиппокампа, гипоталамуса, ретикулярной формации среднего мозга. По мере укрепления рефлекса реакции нейронов идут на убыль, прежде всего в неспецифических областях и подкорковых структурах, а затем в коре. Специализация условных реакций сопровождается уменьшением числа «обучающихся» клеток, что, видимо, отражает процессы концентрации условного возбуждения. Уменьшаются ответы нейронов как в зоне проекции безусловного, так и условного раздражителей. Однако в специфических подкорковых путях проявления условных реакций сохраняются, в то время как они идут на убыль в корковых зонах проекции условного и безусловного раздражителей.
Например, у кроликов при выработке оборонительного условного рефлекса на звук возникшая вначале условная реакция при дальнейшем укреплении рефлекса практически исчезала у 20–30 % нейронов в слуховой и двигательной зонах коры, хотя в подкорковой области слухового пути (внутреннем коленчатом теле) такую затухающую активность обнаруживали лишь 16 % нейронов. При достаточно длительном многократном повторении сочетаний, обеспечивающем высокую прочность вырабатываемого условного рефлекса у кошек, можно было достичь практически полного возвращения импульсных реакций значительной части нейронов на сигнальный раздражитель к тому уровню, когда этот раздражитель был индифферентным (табл. 2).
Таблица 2. Динамика условных реакций 100 нейронов коры мозга кошки при выработке и упрочении зрительно-двигательного условного рефлекса (по А.Б. Когану)
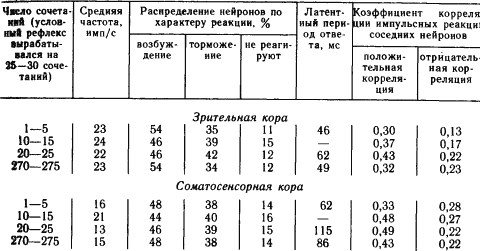
Из табл. 2 видно, что такие показатели условных реакций популяций нейронов, как средняя частота импульсов в ответе и его характер при очень прочном укреплении рефлекса, возвращаются в исходное состояние. От них отстают показатели латентных периодов, имеющие еще высокие значения. Наиболее стабильными оказались условные изменения коэффициентов корреляции импульсных разрядов. Обратное развитие условных реакций именно корковых нейронов при упрочении условного рефлекса можно объяснить концентрацией возбуждения, покидающего исследуемые нейроны, или «передачей» автоматизированного навыка на более низкие уровни управления. Однако сохранение при этом показателей корреляции импульсных потоков, установившихся в результате выработки условного рефлекса, указывает на то, что если при формировании временной связи происходят изменения деятельности обширных популяций нейронов, то после завершения процесса его наличие проявляется главным образом в новых сложившихся при этом межнейронных отношениях.