При отсутствии внешней нагрузки волокно сокращается с максимальной скоростью. При увеличении нагрузки скорость сокращения волокна уменьшается и по достижении определенного уровня нагрузки падает до нуля, при дальнейшем увеличении нагрузки волокно удлиняется.
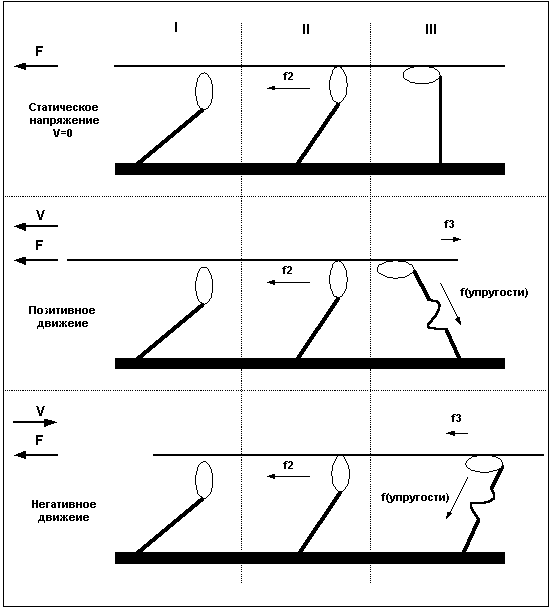
Причину в различии силы волокна при различных направлениях движения легко понять, рассмотрев уже приведенный ранее пример с гребцами и веслами. Дело в том, что после завершения «гребка» миозиновый мостик некоторое время находится в состоянии сцепления с нитью актина, представьте, что весло после гребка так же не сразу извлекается из воды, а находится погруженным еще некоторое время. В случае, когда гребцы плывут вперед (позитивное движение), весла, остающиеся погруженными в воду после завершения гребка, тормозят движение и мешают плыть, в то же время, если лодка буксируется назад, а гребцы сопротивляются этому движению, то погруженные весла так же мешают движению, и буксиру приходится прилагать большие усилия. То есть, при сокращении волокна, сцепленные мостики мешают движению и ослабляют силу волокна, при негативном движении – удлинении мышцы – не отцепленные мостики так же мешают движению, но в этом случае они как бы поддерживают опускающийся вес, что и позволяет волокну развивать большую силу. Легче всего понять различия между статическим напряжением, позитивным и негативным движением взглянув на Рис.5.
Итак, мы рассмотрели основные факторы, влияющие на силу и скорость сокращения отдельного волокна. Сила же сокращения целой мышцы зависит от количества волокон в данный момент времени вовлеченных в работу.
Рис.5
Вовлечение волокон в работу.
При поступлении от ЦНС (центральной нервной системы) к мотонейронам (расположенным в спинном мозге), возбуждающего сигнала (запускающий импульс), мембрана мотонейрона поляризуется, и он генерирует серию импульсов, направляемых по аксону к волокнам. Чем сильнее воздействие на мотонейрон (поляризация мембраны), тем выше частота генерируемого им импульса – от небольшой стартовой частоты (4-5 Гц), до максимально возможной, для данного мотонейрона, частоты (50 Гц и более). Быстрые мотонейроны способны генерировать гораздо более высокочастотный импульс, чем медленные, поэтому сила сокращения быстрых волокон гораздо больше подвержена частотной регуляции, чем сила медленных.
В то же время имеется и обратная связь с мышцей, от которой поступают тормозящие сигналы, уменьшающие поляризацию мембраны мотонейрона и снижающие его ответ.
Каждый мотонейрон имеет свой порог возбудимости. Если сумма возбуждающих и тормозящих сигналов, превышает этот порог и на мембране достигается необходимый уровень поляризации, то мотонейрон вовлекается в работу. Медленные мотонейроны имеют, как правило, низкий порог возбудимости, а быстрые – высокий. Мотонейроны же целой мышцы имеют широкий спектр значений этого параметра. Таким образом, при повышении силы сигнала ЦНС, активируется все большее число мотонейронов, а мотонейроны с низким порогом возбудимости увеличивают частоту генерируемого импульса.
Когда требуется легкое усилие, например, при ходьбе или беге трусцой, активируется небольшое число медленных мотонейронов и соответствующее число медленных волокон, ввиду высокой выносливости этих волокон такая работа может поддерживаться очень долго. По мере увеличения нагрузки ЦНС приходится посылать все более сильный сигнал, и большее число мотонейронов (а, следовательно, волокон), вовлекается в работу, а те, что уже работали, увеличивают силу сокращения, по причине увеличения частоты импульсации, поступающей от мотонейронов. По мере увеличения нагрузки в работу включаются быстрые окислительные волокна, а по достижения определенного уровня нагрузки (20%-25% от максимума), например, во время подъема в гору или финального спурта, силы окислительных волокон становится недостаточно, и посылаемый ЦНС сигнал включает в работу быстрые – гликолитические волокна. Быстрые волокна значительно повышают силу сокращения мышцы, но, в свою очередь, быстро утомляются, и в работу вовлекается все большее их количество. Если уровень внешней нагрузки не уменьшается, работу в скором времени приходится останавливать из-за усталости, в результате накопления молочной кислоты.
Скорость движения мышцы зависит от соотношения внешней нагрузки и количества вовлеченных в движение волокон. Мобилизация относительно небольшого количества волокон, приведет к медленному движению, как только совместная сила сокращающихся волокон превысит уровень внешней силы. Мобилизация все большего количества волокон при той же внешней нагрузке приведет к увеличению скорости сокращения, а увеличение внешней нагрузки, или падение силы волокон в результате усталости, при неизменном количестве мобилизованных волокон, приведет к падению скорости сокращения.
При предельных нагрузках, например, при подъеме максимального веса или подъеме относительно небольшого веса, но с максимальной скоростью, сокращается сразу максимально возможное для данного индивида число волокон.
По вопросу вовлечения волокон при предельных нагрузках в литературе приводятся противоречивые сведения. Так ряд авторов утверждает, что даже у тренированных спортсменов при предельных нагрузках активно не более 85% от общего количества двигательных единиц в мышце. С другой стороны, Гурфинкель и Левин приводят данные, что большая часть двигательных единиц рекрутируется уже при нагрузках до 50% от максимума, и в дальнейшем, при повышении силы, подключается лишь небольшая часть (около 10%) самых крупных двигательных единиц, а рост силы от 75% до 100% происходит уже не за счет вовлечения новых двигательных единиц, а за счет повышения частоты импульса, генерируемого мотонейронами. Независимо от того, чья точка зрения является правильной, важно одно – сила мышцы зависит от силы импульса ЦНС, а чем обусловлен рост силы при нагрузках свыше 75% от максимальной только ли ростом частоты, генерируемой мотонейронами, или еще и вовлечением в работу новых мотонейронов не принципиально. Важно так же и то, что в мышцу встроен ограничительный механизм контроля над развиваемым напряжением, осуществляемый через сухожильные органы Гольджи. Сухожильные рецепторы регистрируют напряжение и при превышении критического значения оказывают тормозящее воздействие на мотонейроны. Считается, что при отключении контроля над напряжением проявляется «сверхсила» человека, регистрируемая в экстремальных ситуациях.