Но не все миобласты идут по пути слияния, часть из них обособляется в виде клеток-сателлитов, располагающихся на поверхности мышечного волокна, а именно в сарколеме, между плазмолемой и базальной мембраной – составными частями сарколемы. Клетки-сателлиты, в отличие от мышечных волокон, не утрачивают способность к делению на протяжении всей жизни, что обеспечивает увеличение мышечной массы волокон и их обновление. Восстановление мышечных волокон при повреждении мышцы возможно благодаря клеткам-сателлитам. При гибели волокна, скрывающиеся в его оболочке, клетки-сателлиты активизируются, делятся и преобразуются в миобласты. Миобласты сливаются друг с другом и образуют новые мышечные волокна, в которых затем начинается сборка миофибрилл. То есть при регенерации полностью повторяются события эмбрионального (внутриутробного) развития мышцы.
Помимо многоядерности отличительной чертой мышечного волокна является наличие в цитоплазме (в мышечном волокне ее принято называть саркоплазмой) тонких волоконец – миофибрилл (рис.1), расположенных вдоль клетки и уложенных параллельно друг другу. Число миофибрилл в волокне достигает двух тысяч. Миофибриллы являются сократительными элементами клетки и обладают способностью уменьшать свою длину при поступлении нервного импульса, стягивая тем самым мышечное волокно. Под микроскопом видно, что миофибрилла имеет поперечную исчерченность – чередующиеся темные и светлые полосы. При сокращении миофибриллы светлые участки уменьшают свою длину и при полном сокращении исчезают вовсе. Для объяснения механизма сокращения миофибриллы около пятидесяти лет назад Хью Хаксли была разработана модель скользящих нитей, затем она нашла подтверждение в экспериментах и сейчас является общепринятой.
Механизм сокращения волокна.
Чередование светлых и темных полос в миофибрильной нити определяется упорядоченным расположением по длине миофибриллы толстых нитей белка миозина и тонких нитей белка актина; толстые нити содержатся только в темных участках (А-диск) (рис.3), светлые участки (I-диск) не содержат толстых нитей, в середине I-диска находится Z-линия – к ней крепятся тонкие нити актина. Участок миофибриллы, состоящий из А-диска (темной полосы) и двух половинок I-дисков (светлых полос), называют саркомером. Сокращение длины саркомера происходит путем втягивания тонких нитей актина между толстыми нитями миозина. Скольжение нитей актина вдоль нитей миозина происходит благодаря наличию у нитей миозина боковых ответвлений, называемых мостиками. Головка миозинового мостика сцепляется с актином и изменяет угол наклона к оси нити, тем самым как бы продвигая нить миозина и актина относительно друг друга, затем отцепляется, сцепляется вновь и вновь совершает движение. Перемещение миозиновых мостиков можно сравнить с гребками весел на галерах. Как перемещение галеры в воде происходит благодаря движению весел, так и скольжение нитей происходит благодаря гребковым движениям мостиков, существенное отличие состоит лишь в том, что движение мостиков асинхронно.
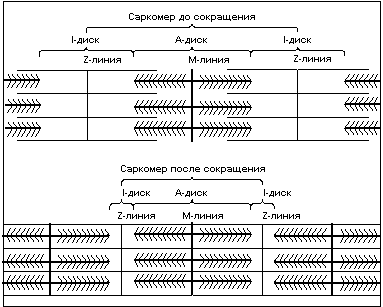
Рис.3
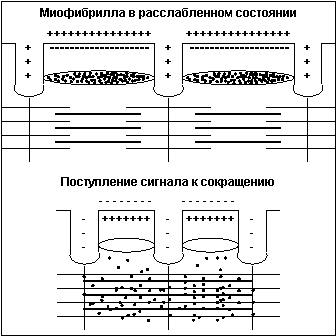
Тонкая нить представляет из себя две спирально скрученные нити белка актина. В канавках спиральной цепочки залегает двойная цепочка другого белка – тропомиозина. В расслабленном состоянии мостики миозина не имеют возможности связаться с актином, так как места сцепления блокированы тропомиозином. При поступлении по аксону двигательного мотонейрона нервного импульса клеточная мембрана меняет полярность заряда, и из специальных терминальных цистерн, расположенных вокруг каждой миофибриллы вдоль всей ее длины, в саркоплазму выбрасываются ионы кальция (Са++) (рис.4).
Рис.4
Под воздействием Са++ нить тропомиозина входит глубже в канавку и освобождает места для сцепления миозина с актином, мостики начинают цикл гребков. Сразу после высвобождения Са++ из терминальных цистерн он начинает закачиваться обратно, концентрация Са++ в саркоплазме падает, тропомиозин выдвигается из канавки и блокирует места сцепления мостиков – волокно расслабляется. Новый импульс опять выбрасывает Са++ в саркоплазму и все повторяется. При достаточной частоте импульсации (не менее 20 Гц) отдельные сокращения почти полностью сливаются, то есть достигается состояние устойчивого сокращения, называемое тетаническим сокращением или гладким тетанусом.
Энергетика мышцы.
Естественно, что для движения мостика требуется энергия. Как я уже упоминал ранее, универсальным источником энергии в живом организме является молекула АТФ. Под действием фермента АТФазы АТФ гидролизуется, отсоединяя фосфатную группу в виде ортофосфорной кислоты (Н3РО4), и превращается в АДФ, при этом высвобождается энергия.
АТФ + H2O = АДФ + H3PO4 + энергия.
Головка миозинового мостика при контакте с актином обладает АТФазной активностью и соответственно возможностью расщеплять АТФ и получать энергию, необходимую для движения.
Запас молекул АТФ в мышце ограничен, поэтому расход энергии при работе мышцы требует постоянного его восполнения. Мышца имеет три источника воспроизводства энергии: расщепление креатинфосфата; гликолиз; окисление органических веществ в митохондриях.
Креатинфосфат обладает способностью отсоединять фосфатную группу и превращаться в креатин, присоединяя фосфатную группу к АДФ, которая превращается в АТФ.
АДФ + креатинфосфат = АТФ + креатин.
Эта реакция получила название – реакции Ломана. Запасы креатинфосфата в волокне не велики, поэтому он используется в качестве источника энергии только на начальном этапе работы мышцы, до момента активизации других более мощных источников – гликолиза и кислородного окисления. По окончании работы мышцы реакция Ломана идет в обратном направлении, и запасы креатинфосфата в течение нескольких минут восстанавливаются.
Гликолиз – процесс распада одной молекулы глюкозы (C6H12O6) на две молекулы молочной кислоты (C3H6O3) с выделением энергии, достаточной для «зарядки» двух молекул АТФ, протекает в саркоплазме под воздействием 10 специальных ферментов.