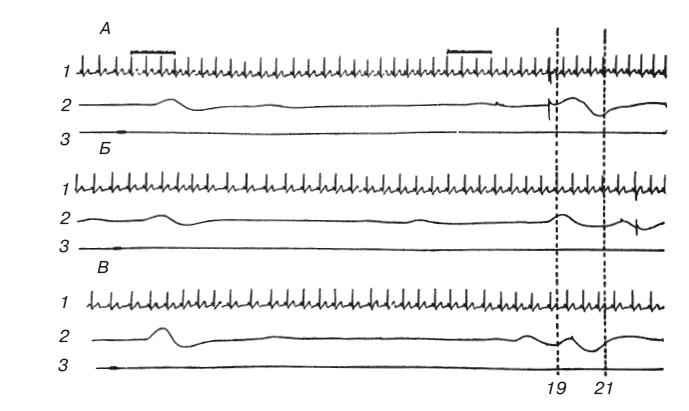
Рис. 1. Условный оборонительный рефлекс у человека
А, Б – ошибочные реакции, сопровождавшиеся болевым раздражением током; В – правильная реакция; 1 – ЭКГ; 2 – КГР, отметка реакции и тока; 3 – звуковой сигнал. Прерывистыми линиями отмечена «зона безопасности»
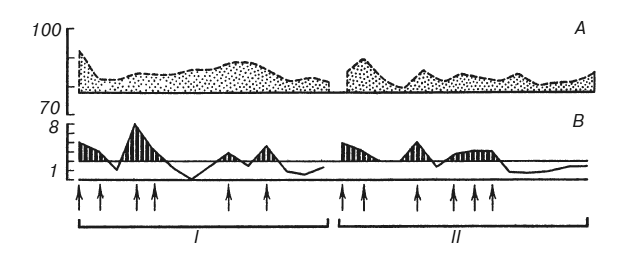
На рис. 2 сплошная кривая показывает время двигательных реакций в последовательных пробах. Раздражения током наносились, когда ошибки субъекта превышали допустимое отклонение (обозначено двумя горизонтальными линиями). Прерывистая линия показывает изменения продолжительности шести сердечных сокращений в пересчете на число ударов в минуту. Верхняя горизонтальная линия обозначает среднюю частоту пульса в фоне до начала выработки условного рефлекса.
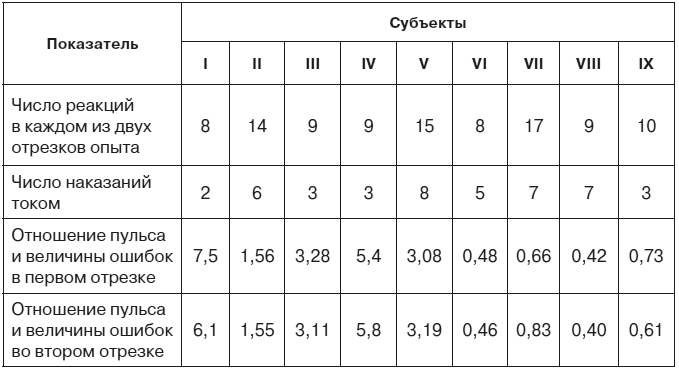
Если сравнить две стадии эксперимента, содержащие одинаковое число проб и одинаковое количество болевых раздражений током, можно убедиться, что суммарное отклонение частоты сердцебиений от исходного фона в этих двух частях эксперимента различно. Следовательно, оно зависит не только от количества наказаний. Опыты на 9 испытуемых показали, что суммарное изменение частоты сердцебиений пропорционально суммарному отклонению времени двигательных реакций от заданной величины, то есть пропорционально степени совершенства, точности и надежности условного оборонительного рефлекса (табл. 1). Это правило справедливо и для тех случаев, где величина ошибок (но не их количество, не количество болевых раздражений) нарастала вместе с нарастанием частоты сердцебиений и где, следовательно, динамику вегетативных сдвигов нельзя было объяснить привыканием к болевым стимулам по мере их повторения.
Рис. 2. Выработка условного оборонительного рефлекса
А — нарастание частоты сердцебиений по сравнению с фоном (уд/мин); В – отклонение времени двигательных реакций от заданной величины (с). Абсцисса – последовательные пробы на протяжении двух стадий эксперимента I и II. Стрелки – раздражения током
Таблица 1. Отношение суммарного изменения частоты сердцебиений к суммарной величине ошибок в опытах с девятью исследуемыми лицами
Эксперименты на животных также показывают, что мозг прогнозирует вероятность наказания в зависимости от степени совершенства инструментального двигательного рефлекса. Л. А. Преображенская вырабатывала условный оборонительный рефлекс у собак в ситуации, где подъем передней лапы до определенного уровня и удержание ее на этом уровне в течение 10 с предотвращали болевое раздражение током противоположной задней лапы (методика Г. П. Зеленого, Г. В. Скипина, Р. Л. Винник и др.). Условный звуковой сигнал подавали за 10 с до болевого раздражения. Сочетание звука с болевым раздражением до выработки условного двигательного рефлекса вело к нарастанию амплитуды и процентного содержания тета-ритма в частотном спектре электрической активности дорзального гиппокампа (рис. 3). Количественный анализ обнаружил позитивную корреляцию изменений суммарного напряжения гиппокампального тета-ритма (измеренного по показаниям интегратора) с частотой сердцебиений. Оба симптома заметно ослабевали по мере стабилизации двигательного навыка, надежно избавляющего животное от боли. Любые затруднения в осуществлении движений вели к повторному нарастанию тета-ритма. Таким образом, опыты Л. А. Преображенской [1969; см. также: Konorski, Santibanez, Beck, 1968] показали, что интенсивность гиппокампального тета-ритма зависит не от двигательной активности самой по себе, но от эффективности двигательных актов, от их влияния на вероятность предотвращения болевых раздражений. Вместе с тем на степень эмоционального напряжения оказывает свое влияние и потребностно-мотивационный фактор: в опытах с оборонительными условными рефлексами суммарное напряжение тета-ритма и учащение сердцебиений были выражены значительно сильнее, чем в опытах с пищевыми.
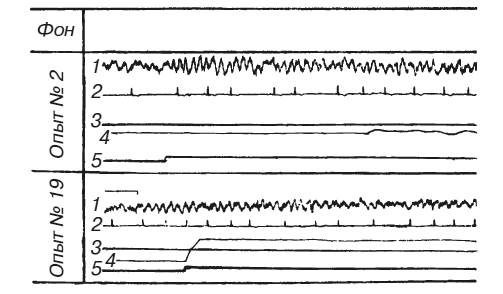
Рис. 3. Изменения гиппокампального тета-ритма и частоты сердцебиений во время выработки инструментального условного оборонительного рефлекса у собаки
1 – ЭГ дорзального гиппокампа; 2 – ЭКГ; 3 – уровень выключения тока; 4 – подъемы передней лапы, выключавшей ток; 5 – условный звуковой сигнал (по Л. А. Преображенской)
Заканчивая обзор данных, свидетельствующих об исчезновении эмоционального напряжения по мере выработки адекватной условной реакции, П. Фресс пишет: «Прежде всего следует подчеркнуть, что не существует эмоциогенной ситуации как таковой. Она зависит от отношения между мотивацией и возможностями субъекта» [Фресс, 1975, с. 133]. Предположение о том, что даже хорошо упроченный инструментальный оборонительный условный рефлекс продолжает мотивироваться страхом, как бы замещающим потребность избегания боли, подвергнуто аргументированной критике Ж. Кюттеном: «…потребность избегания болезненного стимула продолжает действовать (до тех пор, пока животное не усвоит, что сигнал не предвещает опасности), но поскольку животное не испытывает ни боли от стимула, ни угрозы опасной ситуации, оно не проявляет более эмоциональной реакции страха» [Кюттен, 1975, с. 70].
Литература переполнена экспериментальными данными, свидетельствующими о зависимости эмоционального напряжения от величины потребности (мотивации) и прогнозирования вероятности ее удовлетворения. Например, было установлено, что частота пульса у банковских служащих зависит от степени их ответственности (счет банкнотов различного достоинства) и количества информации, содержащейся в одной операции [Gantchev, Danev, Kitcheva, 1967]. Эмоциональные реакции обезьян закономерно зависят от изменения вероятности пищевого подкрепления [Meiges, Popper, 1976]. По данным Д. Н. Меницкого и М. М. Хананашвили [1969], наибольшее эмоциональное напряжение у собак (визг, лай, чесание, царапанье кормушки) наблюдалось при вероятности подкрепления 1:4, а по мере продолжения опыта – при 1:2. Значение информационного фактора выступает особенно отчетливо в опытах со спаренными животными, когда оба партнера получают равное количество ударов током, но только один из них может предотвратить наказание соответствующей инструментальной реакцией. Показано, что именно у этого животного постепенно исчезают признаки страха, предотвращается изъявление слизистой желудка и кишечника [Desiderato, Newman, 1971; Jonas, Jonas, 1975; Starr, Mineka, 1977]. Мотивационный и информационный факторы генеза эмоционального напряжения имеют различный вес у разных животных одного и того же вида. По данным Л. А. Преображенской [1974], максимальное учащение сердечного ритма наблюдалось у одних собак при 30 %-ном пищевом подкреплении, а у других – при 5 %-ном. Следовательно, для собак второй группы суммарный «проигрыш» в удовлетворении пищевой потребности имел большее значение, чем неопределенность экспериментальной ситуации.
Имеются сведения и о том, что две составляющие эмоционального напряжения по-разному сказываются на величине различных вегетативных сдвигов. В опытах с участием человека величина побуждения (размер платы за правильное решение) преимущественно влияла на частоту пульса, дыхания и уровень электрического сопротивления кожи, а трудность задачи (количество выборов) – на объемный пульс и кожногальванический рефлекс [Wilkinson, El-Beheri, Gieseking, 1972]. Преимущественная связь кожногальванического рефлекса с информационным фактором обнаружена и в опытах, где кожногальванический рефлекс был слабее при ожидании болевого раздражения током с высокой вероятностью, чем при более редких, но трудно прогнозируемых ударах [Epstein, Bahm, 1971].