Важно отметить, что хотя оба переносчика выполняют сходные функции в общей мембране митохондрии, однако их работа организована таким образом, что они не мешают друг другу. Убихинон функционирует в толще мембраны, а цитохром С мигрирует по наружной её поверхности. Пространственное разделение обоих потоков позволяет исключить случаи транспортного хаоса.
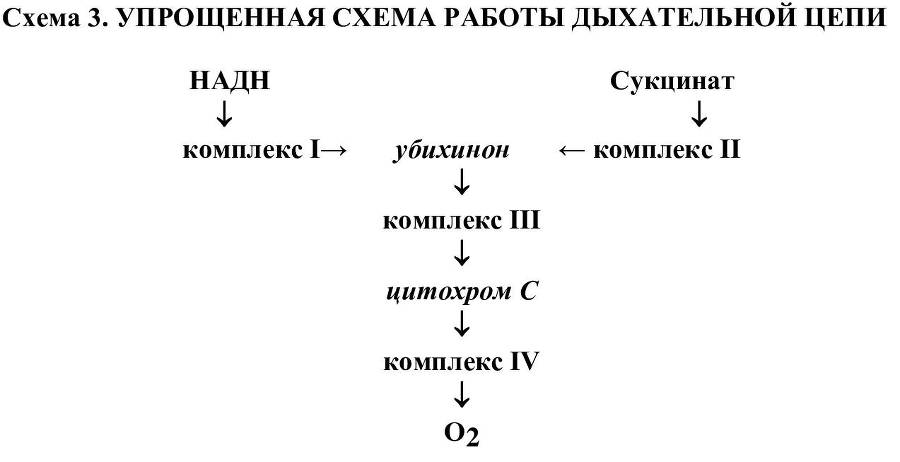
В качестве энергетического субстрата комплекс I использует НАДН (никотинамидадениндинуклеотид), образующийся в процессах как аэробного, так и анаэробного окисления субстратов. Комплекс II катализирует реакции окисления сукцината, образующегося в цикле трикарбоновых кислот (цикл Кребса).
Кислород является субстратом митохондриальной дыхательной цепи. Дефицит кислорода ведет к ограничению и к быстрой дезорганизации работы дыхательной цепи, её мультиферментной системы. Главным результатом в этом случае является истощение клеточных запасов макроэргов и повреждение энергопребразующих механизмов. Такую гипоксию в настоящее время принято называть биоэнергетической. При нарушении энергетических потоков наступает состояние гипоксии тканей.
В зависимости от причин, вызывающих энергетический дефицит, различают 4 вида гипоксии.
Гипоксическая гипоксия возникает при снижении парциального давления кислорода в легочных альвеолах, что чаще всего наблюдается при расстройстве системы внешнего дыхания или, при подъеме на высоту, в горах.
Гемическая гипоксия наступает при дефиците или нарушении функционирования эритроцитарных клеток, специализирующихся на доставке кислорода из легких в ткани-потребители. При этом различают анемический тип гемической гипоксии, связанный с уменьшением концентрации гемоглобина, и патологический тип, наблюдаемый при инактивации гемоглобина.
Циркуляционная гипоксия – это гипоксия тканей связана с нарушением кровообращения. Она может быть связана как с местным, так и системным нарушением кровообращения или перфузии ткани кровью.
Тканевая гипоксия, рассматривается отдельно в случае, когда парциальное давление кислорода соответствует норме, гемоглобин эффективно связывает молекулы кислорода, а кровь успешно доставляет его в ткани, но последние не в состоянии его использовать. Тканевая гипоксия наступает при отравлении дыхательными ядами митохондриальных ферментов, например, цианидами или угарным газом, при дефиците окислительно-восстановительных ферментов или их посредников, возникающем при гормональной недостаточности, а также при повреждении митохондриальных мембран. К тканевой гипоксии как критическому состоянию приводит любой вид неустойчивой гипоксии.
Гипоксию можно рассматривать как одну из разновидностей стрессовых состояний и в соответствии с законом Селье следует ожидать 3 фазы ответа организма на стресс.
На первой фазе – фазе возбуждения – наблюдается усиление активности окисления первичного субстрата и увеличение продукции АТФ. Этой фазе соответствует усиление функциональной активности клеток, в частности интенсификация обменных процессов, в том числе ионного обмена. Происходит мобилизация функций жизненно важных органов. Повышается сократительная активность миокарда, частота сердечных сокращений, артериальное давление. Усиливающуюся гипоксию тканей сердце пытается скомпенсировать увеличением скорости доставки свежих порций крови на периферию. Повышается ответственность за управлением всеми функциями организма со стороны мозга, поэтому растет импульсная активность нейронов, увеличивается частота дыхания. Происходит экстренный выброс гормонов для мобилизации организма к стрессу, в первую очередь для усиления доставки тканям энергетического субстрата. Мобилизуются функции печени для переработки возрастающих потоков недоокисленных метаболитов. Длительность фазы возбуждения определяется продолжительностью гипоксического воздействия и интенсивностью его проявления. Эти процессы определяют суть тренировочного воздействия.
Фаза возбуждения сменяется фазой адаптации. Из-за ограниченности поступления кислорода в ткани происходит постепенное увеличение доли восстановленных форм дыхательных ферментов: и ингибирование комплекса I дыхательной цепи. При этом возрастает доля восстановленной формы убихинона – убихинола. Последний является активатором сукцинатдегидрогеназного комплекса. В результате происходит переключение субстратного участка дыхательной цепи с комплекса I на комплекс II, а в клетке начинают накапливаться НАД-зависимые субстраты цикла Кребса. В этот период времени, несмотря на нарушение работы комплекса I за счет скомпенсированной работы комплекса II, внутриклеточная концентрация АТФ сохраняется неизменной или почти неизменной. При сохранении энергетического гомеостаза функциональная активность клеток также не меняется. Начальный период гипоксии, в течение которого сохраняется состояние энергетического гомеостаза в клетках, относится к компенсированной стадии биоэнергетической гипоксии.
По мере развития гипоксии и снижения запасов кислорода в тканях наблюдается переход к заключительной фазе реакции организма на стресс – фазе истощения. На этой фазе можно выделить два последовательно проходящих этапа деградации электронтранспортных функций митохондрий. На первом этапе наблюдается подавление биоэнергетических функций дыхательной цепи в области комплекса III. Этот период соответствует началу не скомпенсированных изменений и сопровождается снижением содержания макроэргов в клетках.
Нарушение энергетического гомеостаза событие с далеко идущими последствиями для клеток. По этой причине в экстренном порядке мобилизуются внутренние резервы для ликвидации энергетического дефицита. Осуществляется запуск запасной биоэнергетической системы – системы анаэробного окисления субстрата. Происходит централизация кровообращения, при котором не «отключается» от перфузии только сердце, головной мозг и почки – основные жизненно важные системы.
Включение процессов гликолиза происходит в тот момент, когда в клетке снижается содержание АТФ и увеличивается концентрация АДФ и АМФ. Этот факт свидетельствует о прекращении энергетического гомеостаза. Клетка переходит в новое нестабильное состояние и возникает реальная угроза для ее существования. Дальнейшая судьба клетки зависит от энергетических и субстратных потоков, а также от ряда биохимических и биофизических процессов, которые в экстренном порядке запускаются по мере дезэнергизации клетки.
По мере развития гипоксии наблюдается поэтапное повреждение элементов дыхательной цепи. После последовательного подавления переноса электронов через комплексы I, II и III в дыхательной цепи сохраняется последняя возможность образования АТФ за счет работы цитохромоксидазы (комплекса IV). Но, в условиях усиливающейся гипоксии и дезорганизации работы многих ферменных систем, сохранивший работоспособность фрагмент дыхательной цепи уже не способен удовлетворить энергетические запросы клетки. Этому обстоятельству способствует также относительный и абсолютный субстратный дефицит. Как известно, субстратом для комплекса IV является восстановленная форма фермента цитохрома С. Последняя окисляется кислородом с помощью цитохромоксидазы и превращается в окисленную форму фермента. При инактивации комплекса III, в котором обычно осуществлялось ферментативное восстановление окисленной формы фермента, наступает относительный субстратный дефицит. В этих условиях клетка использует запасные механизмы восстановления фермента за счет реакций не ферментативного взаимодействия последнего с убисемихиноном или супероксидным ион-радикалом. Таким образом, удается восстановить поставку субстрата для комплекса IV, нарушенную в результате инактивации в комплексе III.
Однако относительный субстратный дефицит вскоре сменяется на абсолютный. Последнее обстоятельство связано с повреждением мембранных структур. По мере поэтапного выключения в условиях гипоксии отдельных комплексов дыхательной цепи наблюдается последовательное снижение сопрягающих функций митохондрий. Открываются протонные каналы во внутренней мембране митохондрий, что обеспечивает на время поступление свежих партий макроэргов. И за их появление приходится платить все более дорогую цену. В результате набухания митохондрий и дальнейшего увеличения размера пор из матрикса митохондрии в цитоплазму клетки перемещаются различные субстраты и низкомолекулярные белки, включая цитохром С. Потере последнего способствует снижение мембранного потенциала на митохондриальной мембране. Как известно, молекула фермента имеет избыточный положительный заряд и удерживается на внутренней стороне митохондриальной мембраны преимущественно за счет электростатических сил притяжения. Молекулы цитохрома С, мере снижения величины мембранного потенциала, начинают покидать поверхность мембраны и комплекс IV лишается своего субстрата. Дыхательная активность в этом случае полностью подавляется и клетка гибнет. Как правило, на практике происходит сочетание несколько видов гипоксий.