Здесь есть мрачная «ирония судьбы», возможно, определившая многие наклонности человека. В течение двух миллионов лет своей т. н. доисторической животной жизни (т. е. основного биологического периода), нося в себе поразительные потенциалы агрессии, homo вынужден был довольствоваться ролью падальщика, не способного самостоятельно убивать.
По всем меркам зоологии homo достаточно безобиден. Он лишен даже минимального инструментария хищника. У него нет когтей, клыков, скоростных возможностей, ядовитости, мышечной мощи или уникальных особенностей (вроде тех, что есть, к примеру, у пауков). Но есть редкий по силе и комплектности набор основных агрессий.
Что же позволяет с уверенностью говорить об исключительной агрессивности как о видовом отличии homo, изначально ему присущей и неизменной? Ведь о характере и нравах палеоантропов не сохранилось почти никаких свидетельств; лишь скупые археологические указания на вероятную нормативность каннибализма в их стаях, на детритофагию и странную стагнацию вида в течение примерно двух миллионов лет.
Explico.
Такой вывод становится возможен благодаря наличию двух бесспорных факторов.
Первый фактор – это та часть истории человечества, которая была задокументирована, является общеизвестной и прекрасно характеризует вид homo. Формально «коротенький», но неплохо зафиксированный и исследованный период человеческой истории с 5000 г. до н. э., до наших дней вполне может быть образчиком поведения данного вида.
Конечно, история человечества, начиная с шумеров, – это очень размытый, во многом фантазийный, но (в общем и целом) все же документ, свидетельствующий (в первую очередь) о беспрецедентной в зоологии агрессивности вида.
Exempli causa, массовость и постоянность внутривидовых и межвидовых убийств является не просто практикой homo, но и сущностным стержнем его цивилизации. Более того, обязательным, рутинным условием ее развития и успешности.
Но, быть может, исключительная агрессивность – это свойство homo лишь «фиксированной эпохи», некое правило социализации, которое «замешивается» только на внутривидовой и межвидовой крови и через это правило формирует особый тип поведения?
Отнюдь.
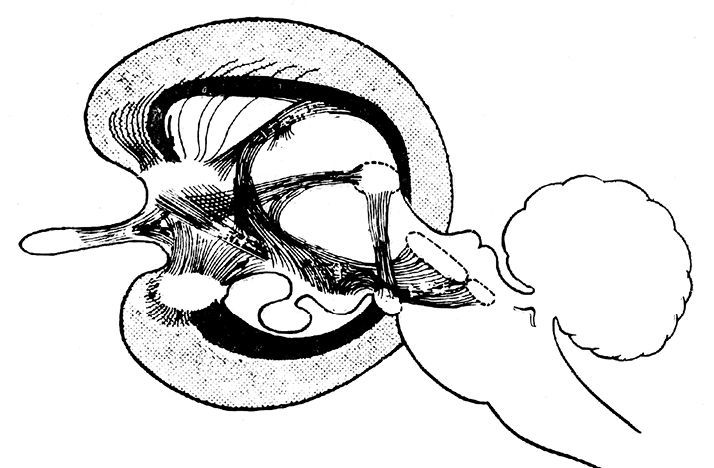
Это предположение легко опровергается знанием как особенностей, так и возраста той структуры головного мозга (лимбической системы systema limbica), которая и генерирует различные эмоции и агрессии, а через них формирует базовое поведение любого позвоночного (Илл. 20).
Илл. 20. Схематическое изображение структур и связи либической системы (по МакЛину)
Еще в 1784 году Поль Брока определил лимбическую систему как область, включающую структуры переднего мозга на границе ростральной части ствола головного мозга у входа в полушария. (Брока же дал ей имя – от limbus, т. е. «кайма».) Впоследствии границы лимбической системы были существенно расширены, но об этом – чуть ниже.
Вглядимся в «фиксированную» историю homo и увидим, что сквозь пламя церковных костров, бесконечье скотобоен, грудей, младенцев, завоеваний, пулеметов, нимбов, концлагерей, шуб, войн, открытий, отбивных, совокуплений, казней и ракет, т. е. сквозь все «знаковые» картинки человеческой цивилизации просвечивают знакомые еще по мозгу древних рептилий очертания амигдалы[22], сосцевидного тела, гипоталамуса, септума и прочих первородных структур systema limbica. (Причем они не просто «просвечивают» и видятся, они во многом и определяют содержание этих картинок.)
Следует помнить, что вышеперечисленные мною отделы мозга окончательно сформировались еще в палеозое, в черепах синапсид (терапсид и пеликозавров), и неизбежно обрекли почти все виды, унаследовавшие их от звероящеров, на агрессивность как на абсолютную норму и основной движитель поведения.
(Лимбическая система мозга функционально и анатомически сходственна у всех видов животных, включая homo. Следовательно, речь может идти о возрасте «вообще» systema limbica, а не конкретно у человека. Понятно, что являясь поздним видом, homo мог только унаследовать те нейрофизиологические структуры, что складывались у позвоночных за сотни миллионов лет.)
Вероятно, следует сразу признать и оговорить, что нейроморфология и до сегодняшнего дня не имеет единого мнения об анатомических границах systema limbica. Ceterum, для нас это не может явиться существенной проблемой, так как объектом нашего внимания являются возраст и функции тех ее структур, принадлежность которых к лимбической системе сомнений не вызывает, это гипоталамус, гиппокамп, миндалевидный (амигдалярный) комплекс, прозрачная перегородка, поясная извилина, вентромедиальные ядра таламуса, центральное серое вещество, мамиллярные тела et cetera).
Наилучшим и наиболее полным образом на сегодняшний день (применительно к эмоциогенным функциям) изучены такие части systema limbica, как гипоталамус и амигдалярный комплекс, в чем можно убедиться на основании следующих трудов: В. Черкес (1967), М. Мгалоблишвили (1974), K. Pribram, Kaplan (1954), Rosvold et al. (1951), Adey (1958), Gloor (1960), Brutkowski, Fonberg, Mempel (1960), Brady et al. (1954), Д. Бирюков (1963), Lissak, Endorczi (1961), Summer, Kaelber (1962), Koikegami (1964), А. Гамбарян и др. (1981), A. Kuntz (1953), Goddard (1964), Nacao (1960), Ursin (1965), Е. Сепп (1959), Kaada (1972), Blanchard (1972), Н. Гращенков (1964), Righetti (1903), Babinski (1900), Economo (1917–1920), Camus, Roussy (1913), П. Анохин (1957), Г. Мэгун (1958), Дж. Папец (1937), W. Cannon, Z. Back, A. Rozenblueth (1931), J. Johnston (1923), W. Le Gros Clark (1936) et cetera.
Для начала, используя данные труды и заключенные в них экспериментальные данные, необходимо расставить точки над i в вопросе «возраста» systema limbica.
Отметим, что мы имеем сумму очень авторитетных мнений по данному поводу, из которых выберем три. (По принципу академического радикализма их авторов.)
I. «Связи меж гипоталамической областью и более каудально расположенными структурами продолговатого и спинного мозга принадлежат к числу ранних» (Гращенков Н. Гипоталамус, 1964).
II. «Гипоталамическая область обнаружена у всех хордовых, в том числе и у тех рыб, что не претерпели существенных эволюционных изменений со времен палеозоя» (К примеру, у латимерии – Прим. автора). (Бирюков Д. Эволюционные идеи И. М. Сеченова и некоторые вопросы физиологии нервной деятельности // Физиологический журнал СССР, 1963).
III. «Миндалевидный комплекс – это симметричная структура, расположенная в вентромедиальной части рострального полюса височной доли. Филогенетически развитые миндалины наблюдаются уже у круглоротых и достигают полного развития у млекопитающих» (Johnston J. Further Contributions to the Study of the Evolution of the Forebrain, 1923).
Легко заметить, что, при всей их несомненности, выводы J. Johnston, Д. Бирюкова, Н. Гращенкова (как и любых других исследователей) излишне лаконичны, постулятивны и нуждаются в комментариях.
Ergo, комментирую.
Первым признаком архаичности церебральной структуры (как известно) является наличие у нее прямых связей со спинным мозгом.
Возьмем для примера эфферентные пути гипоталамуса: медиальный пучок (fasciculus longitudinalis medialis), дорсальный гипоталамо-тектальный пучок (fasciculus longitudinalis dorsalis), обонятельный мезенцефальный пучок, пучок Щютца, сосково-покрышечный пучок Гуддена, сосковидно-таламический пучок et cetera.