Быстрые мышечные волокна (тип II) более крупные и содержат большее количество сократительных элементов и относительно мало миоглобина, поэтому имеют светлую окраску (бледно-розовую). Их называют белыми. Быстрые мышечные волокна развивают большое усилие при сокращении, скорость их сокращения очень высокая, однако в таких волокнах быстро развивается утомление. Быстрые мышечные волокна не могут использовать молочную кислоту как источник энергии. Наоборот, они сами являются источником образования молочной кислоты.
Среди быстрых мышечных волокон выделяют два подтипа: окислительно-гликолитические и гликолитические. Гликолитические быстрые мышечные волокна (подтип II-В) способны к мощной и кратковременной работе. Однако уже через 20–25 с скоростной работы они утрачивают высокую работоспособность. Окислительно-гликолитические быстрые мышечные волокна (подтип II-А) уступают им по силе и скорости сокращения, но могут работать намного больше, используя как аэробные, так и анаэробные механизмы энергообеспечения.
Следовательно, для проявления выносливости при длительной работе необходимо большее число медленных мышечных волокон (тип I). В частности, в мышцах выдающихся стайеров обнаружено до 80 % медленных мышечных волокон. Для проявления выносливости на средних дистанциях желательно иметь большое количество быстрых окислительно-гликолитических мышечных волокон (подтип II-А), а для достижения высоких результатов в спринтерских дисциплинах необходимо соответствующее количество быстрых гликолитических мышечных волокон (подтип II-В). В мышцах выдающихся спринтеров отмечается до 80 % быстрых мышечных волокон.
Характерно, что соотношение волокон разного типа в мышцах определяется наследственными факторами, не зависит от пола и не поддается изменению в процессе спортивной тренировки. С возрастом количество быстрых мышечных волокон постепенно уменьшается. Появившиеся методы «генетического» воздействия на мышечную ткань не меняют тип мышечных волокон, они влияют только на сократительные элементы внутри каждого мышечного волокна.
2. Механизмы обеспечения мышечной работы энергией. Другим важнейшим фактором, определяющим выносливость, является мощность и емкость систем, обеспечивающих энергией мышечную работу.
Единственным источником для мышечных сокращений является АТФ. Её запасов в мышцах хватит всего лишь на 1–2 с интенсивной работы. Однако в мышцах, наряду с распадом АТФ, происходит ее обратный синтез (ресинтез). Ресинтез АТФ в мышцах осуществляется с помощью трех механизмов (энергетических систем):
– фосфагенной (креатинфосфатной);
– гликолитической (лактацидной);
– окислительной (кислородной).
Первые две системы – фосфагенная и гликолитическая – работают по анаэробному пути, третья (окислительная) – по аэробному. Мощность энергетических систем определяется максимальным количеством энергии (молекул АТФ), которое может синтезироваться в единицу времени. Емкость энергетических систем определяется максимальным количеством энергии (молекул АТФ), которое может синтезироваться за все время работы.
Фосфагенная энергетическая система. Известно, что ресинтез АТФ происходит в протоплазме мышечных клеток в непосредственной близи от сократительных волокон (миофибрилл) за счет другого высокоэнергетического соединения – креатинфосфата (КрФ). Конечными продуктами расщепления КрФ являются креатин и свободный фосфат. Данные вещества не снижают работоспособность мышц. Молочная кислота не образуется, поэтому такой механизм называют анаэробно-алактатным (без кислорода и образования молочной кислоты).
Фосфагенная энергетическая система обладает наибольшей мощностью по сравнению с другими системами. Емкость фосфагенной системы невелика, так как запасы АТФ и КрФ в мышцах весьма ограничены. Поэтому фосфагенная система играет решающую роль в энергообеспечении кратковременной работы, осуществляемой с максимально возможными по силе и скорости сокращениями мышц.
Полное восстановление запасов КрФ в мышцах происходит после окончания работы. Для этого требуется 2–5 л кислорода и несколько минут времени (не более 10).
Гликолитическая энергетическая система. В основе функционирования этой системы, обеспечивающей ресинтез АТФ и КрФ, лежит цепь биохимических реакций анаэробного расщепления углеводов, главным образом гликогена, запасенного в мышцах. В результате таких реакций образуются недоокисленные продукты, главным из которых является молочная кислота. Совокупность этих реакций называется гликолизом. Молочная кислота негативно влияет на работоспособность мышц. Такой механизм называют анаэробно-лактатным (без кислорода, но с образованием молочной кислоты).
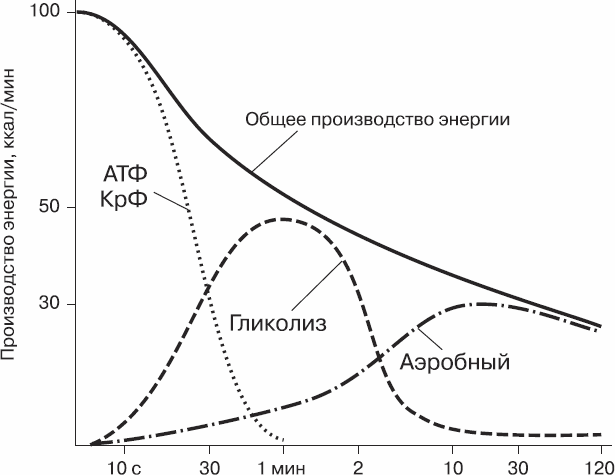
Мощность гликолитической энергетической системы в три раза уступает мощности фосфагенной системы, однако ее емкость в 2–2,5 раза выше, чем емкость последней.
Гликолиз начинается в первые секунды работы, но своего максимума достигает лишь через 30–40 с. Гликолитическая энергетическая система является ведущей при выполнении интенсивной мышечной работы продолжительностью от 20 с до 5 мин.
Полная нейтрализация всей молочной кислоты происходит после окончания работы. Для этого требуется 10–15 л кислорода и до 3 ч времени.
Кислородная энергетическая система. При непрерывном поступлении кислорода в митохондриях мышечных волокон действует кислородная энергетическая система. Для ресинтеза АТФ и КрФ в данном случае используются гликоген, глюкоза и свободные жирные кислоты. В мышечных волокнах имеются небольшие запасы кислорода, связанного с миоглобином. Поэтому для обеспечения работы данной энергетической системы необходимо бесперебойное поступление кислорода в мышечные волокна из крови. Конечными продуктами данной системы являются углекислый газ и вода.
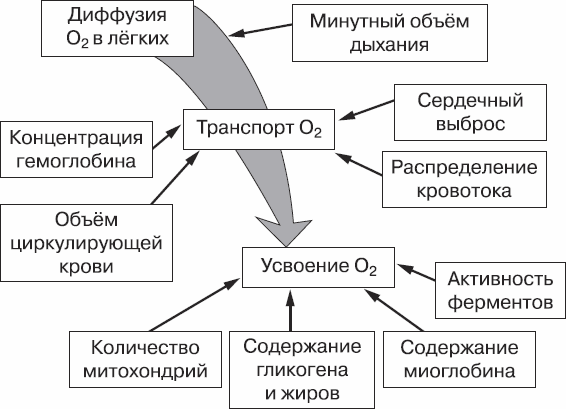
Поступление кислорода в мышечные волокна обеспечивает своеобразный «кислородный конвейер», состоящий из дыхательной и сердечно-сосудистой систем, а также самой крови (рис. 1). Поэтому на эффективность работы кислородной энергетической системы влияют не только запасы энергетических субстратов (углеводов и жиров), но и качество работы многих систем организма. Для оценки работы «кислородного конвейера» чаще всего учитывают величину МПК и способность организма удерживать длительное время потребление кислорода на высоком уровне.
Рис. 1. Схема «кислородного конвейера»
Мощность кислородной энергетической системы уступает мощности фосфагенной системы в 4 раза при окислении углеводов и в 8 раз – при окислении жиров. Емкость кислородной энергетической системы превышает емкость фосфагенной системы в 150–160 раз при окислении углеводов и в 10 000–12 000 раз – при окислении жиров. Поэтому работоспособность мышечного аппарата может поддерживаться кислородной энергетической системой длительное время.
Соотношение трех систем энергообеспечения при мышечной работе показано на рисунке 2.
Рис. 2. Соотношение систем энергообеспечения при мышечной работе
Мощность и емкость энергетических систем повышаются при использовании соответствующих методов тренировки.
3. Кислородный долг. На продолжительность и эффективность физической работы может оказывать влияние наличие и скорость нарастания кислородного долга. Кислородный долг возникает в случае несоответствия кислородного запроса и фактического потребления кислорода во время физической работы.
Кислородный запрос – количество кислорода, необходимое для выполнения работы. В зависимости от мощности физической работы кислородный запрос может составлять от нескольких сот миллилитров до 40 л кислорода в пересчете на минуту. Потребление кислорода у наиболее тренированных спортсменов в зависимости от вида спорта колеблется в пределах от 3,5 до 6 л/мин у мужчин и от 2,5 до 4,5 л/мин у женщин. Только у отдельных выдающихся спортсменов МПК способно превышать эти цифры. Кроме того, потребление кислорода может достигнуть максимума через несколько минут после начала работы.