Частота повреждений ДНК, индуцируемых в каждой клетке организма млекопитающих за 1 час под влиянием фона ионизирующих излучений, составляет примерно 5·10-3. (Оценка сделана экстраполяцией зависимости от дозы числа индуцируемых повреждений ДНК различного типа к фоновым дозам.) Таким образом, скорость возникновения спонтанных повреждений ДНК примерно в 106 раз (а может быть, и в еще большее число раз — мы оценили "вклад" отнюдь не всех механизмов спонтанного повреждения ДНК) превышает скорость возникновения повреждений ДНК, индуцируемых фоновым излучением.
Это заключение может показаться парадоксальным. Ведь генетики, в том числе выдающиеся (например, академик Н. П. Дубинин), считают, что фон излучений вносит хотя и не очень большой, но все же измеримый вклад в частоту спонтанных мутаций у млекопитающих. Как же разрешить этот парадокс?
Механизмы залечивания ДНК и жизнестойкость организмов
Сегодня известно: почти все спонтанные повреждения ДНК залечиваются. Большинство из индуцированных ионизированным излучением повреждений ДНК также репарируется. Однако скорости репарации спонтанных и индуцированных повреждений ДНК могут быть различными. Это станет понятным после того, как мы рассмотрим системы репарации повреждений ДНК. Но сразу же подчеркнем малую исследованность этих систем. Это связано прежде всего с двумя обстоятельствами. Первое можно охарактеризовать вкратце — исследовать процессы репарации спонтанных повреждений трудно из-за ограничений чувствительности традиционных методов исследования. Что касается второго обстоятельства, то для его понимания нужно сделать краткий экскурс в проблему стабильности (или, как теперь становится очевидным, нестабильности) ДНК.
До последнего времени среди молекулярных биологов, биохимиков и особенно генетиков преобладающим являлось мнение о стабильности ДНК. Например, знаменитым Уотсоном в его руководстве по молекулярной биологии подчеркнуто: "Количество ДНК в хромосомах постоянно… В пользу генетической роли ДНК свидетельствовала и ее метаболическая стабильность. В отличие от многих других клеточных молекул ДНК стабильна: атомы, однажды включенные в ее состав, не покидают молекулу".
Однако еще в 1970 году в монографии "Молекулярные механизмы старения" автор этой книги сформулировал концепцию, одно из основных положений которой состояло в том, что в клетках с относительно большой частотой возникают спонтанные изменения генома. Кстати, это заключение было сделано на основании не только анализа свойств ДНК и метаболизма клетки, но и сравнения механизмов старения с механизмами радиобиологических эффектов. Что касается последних, то к тому времени уже были получены факты, позволяющие полагать, что они по крайней мере частично определяются повреждениями ДНК. На основании такого анализа было заключено, что ДНК в клетке должна подвергаться весьма интенсивному повреждению, особенно окислительной деструкции химически активными внутриклеточными метаболитами.
Исходя из этой концепции, а также основываясь на закономерностях, свидетельствующих о роли повреждений ДНК в старении, в лекции, прочитанной в 1972 году в Киеве на IX Международном конгрессе геронтологов, нами было сформулировано следующее следствие: в ряду различных видов млекопитающих существует прямая корреляция между способностью клеток определенного организма к репарации ДНК и видовой продолжительностью жизни.
Несколько лет спустя (в 1974 году) американские радиобиологи Р. В. Харт и Р. Б. Сэтлоу обнаружили такую корреляцию при исследовании репаративного синтеза ДНК, индуцированного УФ излучением в клетках 10 плацентарных млекопитающих, различающихся по видовой продолжительности жизни примерно в 50 раз (от полевки, живущей менее 2 лет, до человека, видовая продолжительность жизни которого составляет около 100 лет).
Однако в этой и последующих работах, в которых также наблюдали корреляцию между продолжительностью жизни организмов и способностью их клеток к репарации ДНК, исследовали репарацию не спонтанных, а индуцированных излучениями повреждений ДНК. И пришли к выводу: часть спонтанных повреждений ДНК устраняется с помощью тех же ферментов, которые участвуют и в репарации повреждений ДНК, индуцируемых УФ или ионизирующим излучением, или химическими мутагенами.
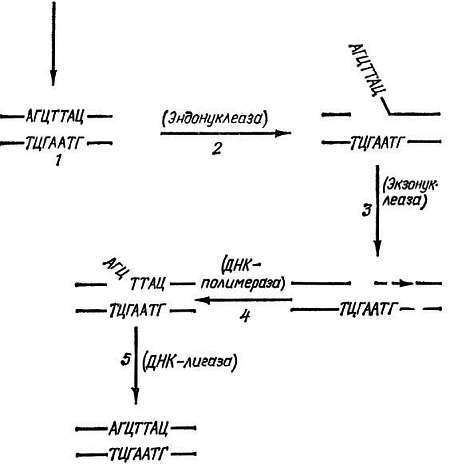
Одна из наиболее изученных систем репарации, защищающих клетки от таких (генотоксических и мутагенных) агентов, состоит из ряда ферментов, залечивающих повреждения в ДНК путем вырезания измененных ее участков и замены их нормальными. Вспомним: хирург, прежде чем зашить концы ран, проводит их хирургическую обработку — удаляет поврежденные участки ткани, "освежает" рану. Процесс репарации начинается с выщепления измененных частей ДНК, например модифицированных оснований. Эта стадия репарации осуществляется также в несколько этапов (рис. 6).
Рис. 6. Схема работы эксцизионных (вырезающих) механизмов репарации ДНК.
1. Индукция повреждения (реакция одного из оснований ДНК с ОН· или Н2О2 или другим эндогенным генотоксическим метаболитом; тепловое повреждение или попадание в ДНК квантов УФ, рентгеновского или другого вида ионизирующего излучения). 2. Узнавание повреждения и образование в ДНК разрыва. 3. Выщепление (вырезание) здорового участка ДНК, прилежащего к поврежденному. 4. Репаративный синтез ДНК. 5. Сшивание концов ДНК. В скобках указаны ферменты, катализирующие соответствующий этап репарации ДНК
Первый из них — "узнавание" повреждения в ДНК и образование вблизи него "надреза" (мы продолжаем проводить аналогию процесса репарации с работой хирурга). Такую операцию осуществляет фермент эндонуклеаза, разрушающий модифицированную нить ДНК. Затем подключается другой фермент — экзонуклеаза, который выщепляет модифицированное основание, а вместе с ним обычно и значительное количество "здоровых" оснований. В результате в ДНК образуется брешь. Поскольку же вторая нить остается целой, то, используя ее как матрицу, третий фермент — ДНК — полимераза заполняет эту брешь. Процесс репарации завершает четвертый фермент — лигаза. Этот фермент связывает вновь синтезированный участок ДНК с тем концом, который остался после процесса выщепления.
Рассмотренный пример наглядно показывает: в процессе репарации происходит синтез ДНК. Этот синтез связан не с делением клетки, а с восстановлением ДНК, поэтому его называют репаративным. Интенсивность его легко определить, добавляя в среду, где находятся клетки, меченый тимидин — вещество, которое входит в состав только ДНК. Теперь, чтобы ответить на вопрос о том, происходил ли "ремонт" ДНК, т. е. замещение отдельных ее оснований, достаточно установить, является ли ДНК клетки, не приступающей к делению, радиоактивной.
С помощью такого сравнительно простого приема нами и было получено доказательство того, что репарация ДНК происходит не только в том случае, когда клетки повреждаются физическими или химическими агентами, т. е. в необычных условиях, но и в таких условиях, которые можно считать нормальными, физиологическими. Поэтому такой процесс получил название спонтанного репаративного синтеза ДНК.
Кстати, вывод, который неизбежно следует из факта протекания спонтанного репаративного синтеза, состоит в том, что ДНК клетки в процессе нормального метаболизма повреждается. Следовательно, биологическая стабильность генома, его надежность обеспечиваются и "волшебной палочкой квантовой химии", т. е. стабильностью ее химических связей (Э. Шредингер), и системой репарации ДНК, функционирующей в клетке.
Только что рассмотренную систему репарации ДНК (по причинам, теперь, наверное, понятным читателю) называют вырезающей или эксцизионной (от англ. exision — вырезать). Одна из принципиальных особенностей репаративного синтеза ДНК, протекающего при такой репарации, состоит в том, что он осуществляется в различные фазы клеточного цикла. "Обычный" же синтез ДНК, в результате которого происходит удвоение количества ДНК и поэтому называемый редупликативным (обычно говорят "репликативный"), протекает лишь в определенную фазу жизни клетки. Таким образом, редупликативный синтез ДНК является запрограммированным, протекающим в S-фазу клеточного цикла, а репаративный — незапрограммированным, или внеплановым.