Эти данные, кроме того, что они подтверждают грустные замечания великих поэтов ("старость — есть второе детство" или "хладея, мы движемся к началу своему"), интересны еще и потому, что приводят к принципиально новому заключению о физиологической роли денатурированных участков ДНК и(или) антител к ним.
Если исходить из традиционной точки зрения (которую для простоты изложения мы рассматривали до сих пор), то нарушения во вторичной структуре ДНК должны возникать лишь случайно и накопление их с возрастом должно происходить постепенно. Но тогда количество денатурированных участков ДНК (и следовательно, концентрация антител к ним) в сыворотке взрослых людей должно было бы быть больше, чем в сыворотке детей. Наблюдается же обратная закономерность. Правда, в своих рассуждениях мы предполагали, что количество антител в сыворотке крови определяется в основном количеством денатурированной ДНК в клетках. Но это может быть и не так. Например, частичная денатурация ДНК может происходить и в сыворотке. А главное, количество синтезируемых антител может и не изменяться параллельно с изменением количества такой ДНК.
Таким образом, анализ возможных изменении с возрастом количества денатурированной ДНК не привел нас к однозначному заключению о характере этих изменений. Но при этом неожиданно приоткрылась, возможно, важная функция такой ДНК и(или) антител к ней. Однако, как уже отмечалось, нативная ДНК может не только денатурировать, но и менять свою конформацию, оставаясь двойной спиралью.
Конформационные изменения ДНК
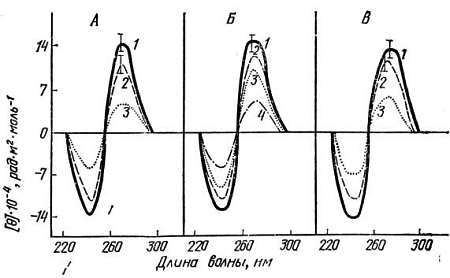
Метод кругового дихроизма (КД) чувствителен к изменениям структуры ДНК такого рода, поэтому В. М. Лобачев, Т. М. Третьяк, А. М. Кузин и автор этих строк его использовали для ответа на вопрос: изменяется ли конформация двойной спирали ДНК в процессе старения? ДНК, выделенная из печени и мозга очень старых крыс (возраст 38 месяцев), имеет спектры, практически идентичные спектрам КД тех же ДНК, облученных ионизирующей радиацией (рис. 9). Оптические активности положительной и отрицательной полос спектров КД ДНК, выделенной из тканей старых животных, или ДНК, облученной в дозе 200 гр, были снижены примерно на 20 % по сравнению с ДНК молодых животных, и это различие было статистически достоверно.
Рис. 9. Спектры кругового дихроизма (КД) ДНК, выделенной из тканей молодых и старых крыс, в сравнении с КД ДНК тимуса теленка.
А. 1. ДНК мозга молодых (возраст 3 мес.) крыс. 2. ДНК мозга старых (возраст 38 мес.) крыс. 3. Облученная ДНК мозга молодых крыс (доза — 1000 ГР).
Б. 1. ДНК печени молодых крыс. 2. Облученная ДНК печени молодых крыс (доза 200 Гр). 3. Облученная ДНК печени молодых крыс (доза 300 Гр). 4. Облученная ДНК печени молодых крыс (доза 1000 Гр).
В. 1. ДНК тимуса теленка. 2. ДНК печени старых крыс. 3. Облученная ДНК печени старых крыс (доза 1000 Гр). (Из работы М. М. Виленчика, Т. М. Третьяк, В. М. Лобачева, А. М. Кузина, Доклады АН СССР, 1981.)
Анализ обнаруженных изменений показал, что наблюденные изменения спектров КД ДНК при старении или после гамма-облучения ДНК не определяются образованием однонитевых разрывов в ДНК; их нельзя также полностью объяснить денатурационными изменениями. Эти изменения спектров КД можно объяснить переходом модифицированных участков ДНК из "обычной" (канонической) В-формы в иную конформацию. Подчеркнем еще раз, что эти конформационные изменения отличаются от ранее изученных денатурационных изменений ДНК.
В то время, когда мы получили первые данные о возрастных изменениях спектров КД, был опубликован ряд работ, в которых было показано, что двуспиральные синтетические полинуклеотиды с определенной последовательностью оснований могут находиться в левоспиральной конфигурации (названной Z-формой), причем спектр КД таких полинуклеотидов оказывается инвертированным (обратным по знаку). Таким образом, к предположению о существовании таких особых форм ДНК разные группы исследователей пришли независимо, исходя из результатов изучения изменений ДНК при старении и анализа физических свойств полинуклеотидов с определенной последовательностью оснований.
Такие последовательности встречаются и в природной ДНК, в частности в ДНК млекопитающих. Вероятно, эти последовательности in vivo со временем также могут переходить в левоспиральные участки, и количество таких участков может возрастать по нескольким причинам. Во-первых, вследствие метилирования оснований, облегчающих переход отдельных участков ДНК из канонической В-конформации в левоспиральную Z-конформацию. Во-вторых, вследствие локального изменения (увеличения) ионной силы в отдельных участках хроматина или накопления в них определенных веществ, также облегчающих такой переход. В-третьих, образованию, а главное "фиксации" изменений конформации определенных участков ДНК in vivo должно способствовать образование в этих участках повреждений первичной структуры, о которых речь шла ранее, а также ковалентных сшивок ДНК — белок и особенно сшивок ДНК — белок-ДНК.
Однако в клетке, вероятно, существуют белки, способные переводить ДНК из левоспиральной в обычную — В-конформацию. И поскольку предполагалось образование левоспиральных участков ДНК после облучения и были основания считать такое образование одним из механизмов повреждающего действия излучения на клетки, то теоретически был получен ответ и на вопрос: в каких именно клетках белки, "репарирующие" левоспиральные участки, нужно искать в первую очередь. Ясно, что в тех, которые очень устойчивы к излучению, в частности, потому, что содержат относительно большое количество "Z-репарирующих белков".
Это предположение было опубликовано в 1981 году, а в конце 1985 года поступило сообщение о том, что в одном из видов бактерий М. Radiodurans, выделенном лет двадцать назад из котлов ядерных реакторов и, следовательно, обладающем исключительно высокой радиоустойчивостью (отсюда и его латинское название), содержатся белки, под влиянием которых ДНК из Z-конформации может возвращаться в обычную В-конформацию.
Раз уж мы коснулись проблемы биологической роли участков ДНК, находящихся в левоспиральной конформации, то отметим, что химические канцерогены также могут индуцировать в B→Z переход. А такие переходы имеют значение в канцерогенезе. Но если это так, то происходящие при старении или после облучения изменения конформации ДНК также могут иметь значение соответственно в спонтанном и радиационном канцерогенезах. Во всяком случае, логичен вопрос: не являются ли B→Z переходы "почвой", подготавливающей развитие рака в пожилом и старческом возрасте? И далее — не обладают ли антиканцерогенными свойствами белки, осуществляющие обратный Z→B переход, т. е. не могут ли они задерживать развитие спонтанного, или химического, или радиационного канцерогенеза? Сформулированные вопросы, как говорится, не только академические: они представляют интерес и в практическом плане. И вполне поддаются исследованию с помощью современных методов, хотя такие исследования должны включать и сложные методики.
Образование "дополнительных" участков ДНК, находящихся в Z-конформации, представляет потенциальную канцерогенную опасность прежде всего потому, что при этом должна нарушиться регуляция функций генома. Имеются данные о роли таких конформации в регуляции активности генов и, возможно, в дифференцировке клетки. А ведь нарушение регуляции генов и состояния дифференцировки клетки многие биологи считают основой канцерогенеза.
Заключение о накоплении при старении клеток участков их ДНК, находящихся в необычных конформациях, не противоречит предположению об их репарируемости. Часть ДНК так плотно упакована в хроматине, что ее измененные участки просто могут быть недоступны для репарирующих ферментов. А если репарирующие ферменты способны находить и репарировать измененные участки ДНК, то по тем или иным причинам осуществляется "залечивание" не всех из них (т. е. могут быть кинетические и термодинамические ограничения).