Глава эта будет построена таким образом. Мы будем последовательно двигаться от момента оплодотворения до полного завершения развития. Проследим за основными этапами формирования кота как трехмерной структуры, состоящей из сотен тканей, которые, в свою очередь, построены из миллионов клеток. Особо выделим мутации, которые нарушают тот или иной этап развития. Таким образом, мы составим представление о том, как организован процесс развития, развертывания генетической информации, и опишем те мутации, которые известны у котов в настоящее время.
Здесь и далее слова «мутация» и «аллель» употребляются как синонимы. По правилам номенклатуры мы будем обозначать доминантные аллели прописными буквами, а рецессивные — строчными. В тех случаях, когда для гена известно более двух аллельных форм, применяются буквенные индексы.
Итак, процесс развития начинается с оплодотворения. Сливаются ядра сперматозоида и яйцеклетки. Оплодотворение — необходимое условие нормального развития. Возможность развития без оплодотворения мы рассмотрим в отдельной главе, посвященной проблеме непорочного зачатия.
Слияние гаплоидных ядер приводит к образованию диплоидной зиготы. Зигота имеет полный набор генов, необходимых для развития. Ядро зиготы последовательно делится митотическим делением на 2,4, 8 и т. д. ядер. Этот период жизни зиготы называется дроблением. Он завершается образованием морулы — группы клеток, похожей на ягоду малины. Клетки на этой стадии еще'мало специализированы. Их можно менять местами, переносить от одного зародыша к другому, смешивать клетки от разных зародышей без видимого влияния на последующее развитие.
Деления клеток продолжаются. Морула превращается в бластоцисту. В ней будущая судьба клеток уже оказывается предопределенной их положением. Клетки, образующие поверхность бластоцисты, затем будут использованы для построения плаценты. Внутренняя клеточная масса даст начало зародышевой оболочке — амниону и желточному мешку, а из остальных клеток разовьется уже собственно зародыш. Из клеток наружного слоя — эктодермы — образуются нервная система и кожные покровы, включая и шерсть. Внутренний слой — эндодерма — дает начало внутренним органам, например пищеварительному тракту. Из мезодермы — среднего слоя клеток — образуются скелет, мышцы, сердце, печень, почки. Далее дифференцировка все более и более углубляется. Выделяются группы стволовых, инициаторных клеток, которые специализируются на создании определенных органов.
Понятно, что все эти события происходят под контролем определенных генов. Отдельные гены и генные ансамбли согласованно включаются в работу и выключаются. Их продукты активируют одни гены и инактивируют другие. Причем в каждой группе клеток играет свой, специфичный только для нее, генный оркестр, и свою, характерную только для нее мелодию.
На ранних этапах развитие у самцов и самок идет практически одинаково. Хотя стартовые позиции в развитии двух полов разные. Вы помните, что самки имеют две Х-хромосомы, самцы только одну.
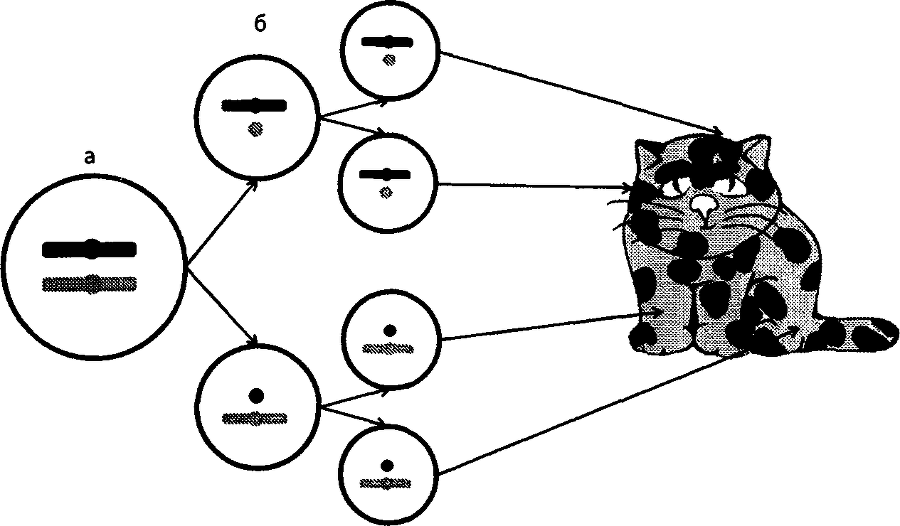
В зиготе (а) активны обе Х-хромосомы. В ходе дифференцировки в ннициаторных клетках (6) одна из Х-хромосом — мутантная (светлая) или нормальная (темная) — инактивируется. Инактивированная хромосома обозначена кружком. Потомки иннциаторных клеток строго наследуют инактивированное состояние Х-хромосомы (в). В тех клетках, где инактивирована нормальная хромосома, проявляется мутантный аллель, и наоборот. В результате формируется мозаичная или черепаховая окраска.
Эта неравноценность устраняется в момент дифференцировки стволовых клеток. На этом этапе одна из двух Х-хромосом у самок инактивируется. То есть она перестает быть доступной для считывания с нее генетической информации. Выбор хромосомы, которая претерпевает инактивацию в момент дифференцировки, определяет случай. В одной стволовой клетке оказывается инактивированной Х-хромосома, доставшаяся от отца, в другой — та, что получена от матери. Но дальше в ряду клеточных поколений это инактивированное состояние одной хромосомы и соответственно активное состояние другой строго наследуются. Иными словами, если в стволовой клетке была инактивирована отцовская Х-хромосома, то во всех потомках этой клетки, в том клоне клеток, который от нее происходит, будет инактивирована именно отцовская Х-хромосома. Таким образом, организм самки можно представить как мозаику клеток: в одних функционирует отцовская Х-хромосома, в других — материнская.
Сколько будет клеток одного или другого типа, решает случай. Судите сами: для каждой клетки вероятность того, что у нее будет инактивирована отцовская Х-хромосома, равна 1/г- Если группа стволовых клеток, нацеленных на создание данного типа тканей, велика, то по закону больших чисел мы можем ожидать, что половина всех клеток в этой ткани у взрослого животного будет иметь инактивированной отцовскую хромосому, а другая половина — материнскую. В том же случае, если эта группа малочисленна — в пределах десятка клеток, — возможны самые
разные соотношения типов клеток во взрослом организме. Может быть даже такая ситуация, что во всех клетках данной ткани будет инактивирована одна и та же Х-хромосома, допустим, материнская.
Каково будет взаимное расположение клеток, принадлежащих к одному клону? Будут ли они располагаться тесной кучкой или смешаются с представителями других клонов? Здесь все зависит от того, о какой ткани идет речь. Клетки некоторых тканей вынуждены мигрировать довольно далеко от того места, где они формируются. В таких случаях перемешивание становится более вероятным.
Наличие разных клеточных клонов у самок очень ярко выявляется на гетерозиготах по мутациям генов, локализованных в Х-хромосоме. У кошек такая мутация есть. Это мутация рыжей окраски. Ее номенклатурное название Orange (символ О).
Проследим за судьбой зародыша женского пола. Он имеет две Х-хромосомы. В одной из них, полученной, например, от отца, находится ген рыжей окраски О, а в другой, полученной от матери, его нормальный аллель о. В момент дифференцировки клеток, потомки которых затем будут заняты синтезом пигментов, в каждой из них случайно инактивируется или одна, или другая X- хромосома. Это довольно небольшая группа клеток: около двухтрех десятков. Но они дают начало тысячам пигментных клеток — меланоцитов. Каждый из меланоцитов будет иметь в инактивированном состоянии именно ту из Х-хромосом, которая инактивировалась в стволовой клетке-родоначальнице.
Меланоциты, в которых активна хромосома, имеющая мутантный аллель, будут синтезировать только желтый пигмент. Меланоциты, несущие в активном состоянии хромосому с нормальным аллелем, произведут оба пигмента, желтый и черный. Волоски, которые снабжаются пигментами из меланоцитов первого типа, будут рыжими, а остальные — серыми. Вся кошка в итоге будет мозаичной, или черепаховой.
Соотношение размеров и степень перемешивания серых и рыжих пятен чрезвычайно разнообразны. Здесь все строится опять-таки на игре случая.
Объясняя феномен черепаховой окраски, мы шли от механизма инактивации. Наука же двигалась обратным путем. Механизм инактивации был придуман для объяснения феномена черепаховой окраски. Затем он был подтвержден целым рядом генетических, цитологических и биохимических экспериментов. Честь открытия этого механизма принадлежит английской исследовательнице Мэри Лайон. Поэтому в научной литературе инактивацию одной из Х-хромосом у самок называют лайонизацией.
Вернемся, однако, к судьбе кошачьего зародыша, которого мы бросили на стадии закладки стволовых клеток.
Зачаток нервной системы — нервная трубка — формируется из парных складок, которые возникают на спинной стороне эмбриона. Из нее затем образуется головной и спинной мозг с периферической нервной системой.
Рассмотрим кратко мутации, которые нарушают развитие нервной системы. Их довольно много. И это понятно. Нормальное
функционирование нервной системы более, чем любой другой системы организма, зависит от точности взаимодействия составляющих ее элементов. Самые незначительные дефекты в функции генов, которые активны даже в локальной группе нейронов, могут приводить к существенным нарушениям поведения.