Рассмотрим теперь несколько форм, наиболее далеких от виноградной улитки и представляющих два других подкласса — переднежаберных и заднежаберных.
Из заднежаберных имеются данные для двух видов голожаберных слизней — тритонии и дендронота. В педальных ганглиях тритонии М. С. Манохина и Л. В. Кузьмина не нашли катехоламиновых нейронов [37]. Также не нашли их и мы в педальных ганглиях дендронота. В отличие от тритонии, у дендронота мы смогли исследовать не только ЦНС, но и периферию. Как в ноге, так и в её деривате — стенке пениса имеется чрезвычайно богатая сеть варикозных катехоламиновых волокон, контактирующих с мышечными клетками. Эти волокна являются отростками клеток, которые расположены в самих этих органах. Каждая катехоламиновая клетка посылает также гладкий, неварикозный отросток в ЦНС, где такие отростки дают начало варикозному синаптическому нейропилю.
Лежащие на периферии педальные катехоламиновые нейроны дендронота были подвергнуты дополнительному электронномикроскопическому исследованию. В подтверждение данных люминесцентной гистохимии было найдено, что в пенисе эти клетки всегда расположены у внутреннего края мышечной стенки, они мелкие, биполярные. Один отросток идет между мышечными клетками в периферическом направлении. Он древовидно ветвится; по ходу ветвей имеются типичные пресинаптические аксональные расширения, контактирующие с клетками второго (снаружи) мышечного слоя. Эти расширения наполнены секреторными гранулами, которые (типично для катехоламинов) имеют плотное зерно при глутаральдегидной префиксации, но лишены его при фиксации одной лишь четырёхокисью осмия. Никогда не наблюдаются контакты между этими аксонами и мышечными клетками наружного слоя, которые вообще не иннервированы. С противоположной стороны от тела нейрона отходит другой отросток, который вступает в состав нервного стволика, идущего между стенкой пениса и семепроводом. Очевидно, это нерв, следующий в ЦНС. На теле катехоламинового нейрона нередко можно видеть синаптические окончания.
Обратимся теперь к переднежаберным моллюскам. Литературных данных о педальных катехоламиновых нейронах для этого подкласса нет, нами исследованы три вида.
У примитивной акмеи Testudinalia (Acmaea) tessellata, не имеющей педальный ганглиев, катехоламиновые нейроны лежат в ноге на небольшом расстоянии от эпителия. Эти клетки биполярные: наружный отросток, лишённый варикозных утолщений, ветвится под клетками эпителия; внутренний направляется к находящемуся в толще ноги педальному ганглиозному стволу, вступая по пути и в самом стволе в состав варикозного катехоламинового синаптического нейропиля. В самих парных ганглиозных педальных стволах, богатых катехоламиновыми нервными окончаниями, отсутствуют нейроны, содержащие катехоламины.
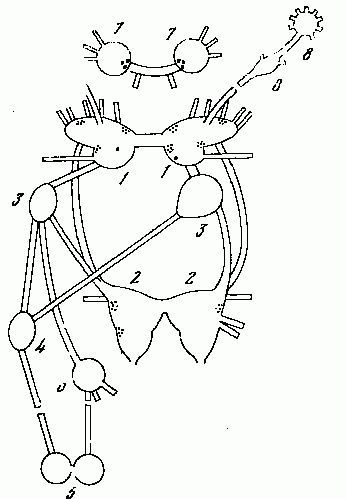
Рис. 14. Центральная нервная система переднежаберного моллюска Pomatiaselegansи позиции центральных нейронов, содержащих биогенные амины. Обозначение ганглиев: 1 — церебральный, 2 — педальный, 3 — плевральный, 4 — супраинтестинальный, 5 — висцеральный, 6 — субинтестинальный, 7 — буккальный, 8 — тентакулярный. В ганглиях 4 — 6 выявление биогенных аминов не проводили. В плевральных и тентакулярных ганглиях моноаминергические нейроны не обнаружены.
Другой исследованный вид — аквариальная «ампуллярия», которая, по сообщению А. А. Шилейко, представляет собой Pila (globosa?) из сем. Pilidae. Улитки этого семейства относятся к группе Architaenioglossa, для которой также характерно отсутствие педальных ганглиев. Парные педальные ганглиозные тяжи связаны многочисленными комиссурами. Такое строение педальной нервной системы рассматривается как древний признак.
В главных педальных стволах и в дочерних тяжах, доходящих почти до подошвы, богато представлены участки синаптического нейропиля с варикозными катехоламиновыми волокнами. Тела катехоламиновых нейронов встречаются в составе ганглиозных стволов. В стенке пениса тела этих нейронов расположены более периферически. Нейроны биполярны, причём периферический отросток не только доходит до эпителия, но и даёт веточки между эпителиальными клетками. Внутренний отросток следует к островкам синаптического нейропиля, расположенным тут же, в стенке органа.
Наконец, третий вид — Pomatias elegans (сем. Pomatiasidae) относится к группе Taenioglossa, представители которой имеют компактные педальные ганглии. В составе этих ганглиев, наряду с большинством нейронов, лишённых биогенных аминов, имеются латерально расположенные группки мелких катехоламиновых клеток (рис. 14). Нейропиль ганглиев богат варикозными катехоламиновыми волокнами. В ноге тоже имеются мелкие катехоламиновые нейроны. Их отростки не подходят к эпителию подошвы. В стенке пениса, которая у этой улитки не является дериватом педальной мышцы, а вырастает из спинной области, катехоламиновых волокон нет совсем.
Эти сравнительные данные позволяют реконструировать картину эволюции катехоламиновых нейронов, входящих в состав педальной нервной системы. Вряд ли можно сомневаться, что их исходная позиция представлена у примитивной акмеи, где эти клетки занимают субэпителиальное положение в подошве ноги. Биполярные эпителиальные или субэпителиальные нейроны обычно рассматриваются как нейроны с сензорной функцией, которую выполняет периферический отросток. Отростки педальных катехоламиновых нейронов проявляют, однако, три рода функциональных отношений. Неварикозный отросток, расположенный под эпителием или входящий в него, очевидно, выполняет сензорные функции; варикозный периферический или центральный отросток, контактирующий с мышечными клетками, мы вправе считать моторным; наконец, варикозный отросток, контактирующий с другими нейронами, служит для осуществления межнейронных связей. Все три рода отношений можно найти в разных группах гастропод. По-видимому, педальные катехоламиновые нейроны, исходно мультифункциональные, в ходе эволюции приобретали разную специализацию, став сензорными в одних случаях, моторными — в других (например, в пенисе дендронота), интернейронами — в третьих.
Непарный дофаминергический интернейрон заслуживает особого упоминания. Берри и Коттрелл [92] исследовали синаптические эффекты этого гигантского нейрона в ЦНС катушки, где он расположен в левом педальном ганглии (ганглий ошибочно обозначен правым в работе [238]). Сравнительно-гистохимические данные, изложенные выше, показывают, что гомологи этой клетки легко прослеживаются в ряду пульмонат. По моим наблюдениям, у прудовика этот нейрон особенно велик и его легко найти в правом педальном ганглии (гомологичном левому ганглию планорбид) рядом с плевро-педальным коннективом; крупные размеры нейрон имеет у слизня, но у хелицид он невелик.
В разных подклассах гастропод педальные катехоламиновые нейроны по-разному вовлечены в процесс ганглионизации. Налицо какая-то избирательность этого процесса, но природа её неясна, хотя общая тенденция прослеживается чётко. Так, среди лёгочных только в примитивном семействе Strophocheilidae нейроны этого типа не обнаружены в составе педальных ганглиев. То, что такие нейроны остались на периферии и у голожаберных, возможно, объясняется их моторными функциями.
Теперь можно ответить на вопрос о том, каким способом катехоламиновые нейроны появились в составе нейронной популяции педальных ганглиев виноградной улитки. Сравнительные данные позволяют с уверенностью утверждать, что клетки с этим типом химической специфичности не возникли в процессе эволюционного дифференцирования исходно однородной нейронной популяции ганглиев: они как специфичная группа существовали до возникновения ганглиев. В процессе дивергентной эволюции гастропод эти клетки ганглионизировались или оставались на периферии; одновременно они приобретали разную функциональную специализацию, что, однако, не сказывалось на их медиаторном химизме.
5. 3. 5. Другие примеры клеточных гомологий