Конечно, физиологические механизмы действия ЭМП не ограничиваются вегетативной нервной системой. Можно раздельно рассматривать чувствительность к ЭМП отдельных участков рефлекторной дуги: рецептор — центральное звено — эффектор. И здесь центральное звено оказалось более уязвимым.
При электрическом раздражении седалищного нерва крысы (находящейся под наркозом) можно регистрировать как биоэлектрические процессы в иннервируемой мышце, так и биопотенциалы различных отделов головного мозга. Если крысу поместить на 10—20 мин в постоянное магнитное поле с индукцией 400 мТ, то в икроножной мышце нельзя было заметить изменений потенциала действия (рис. 16), в то же время вызванный потенциал на электрическую стимуляцию указанного нерва увеличивался по амплитуде и приобретал дополнительные колебания. Сходную картину наблюдали как в сенсомоторной области коры больших полушарий головного мозга, так и в гипоталамусе и в коре мозжечка. Подобные изменения вызванного потенциала наблюдали при повышении уровня бодрствования животного или при увеличении силы раздражения. Иными словами, МП указанных параметров как-то усиливало ответ мозга на электрическую стимуляцию нерва, тогда как на ответ мышцы то же поле влияния не оказывало.
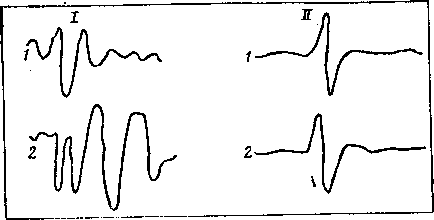
Рис. 16. Вызванный потенциал сенсомоторной коры больших полушарий мозга (I) крысы и потенциал действия икроножной мышцы (II) лягушки на электрическую стимуляцию седалищного нерва в фоне (1) и после 10—20-минутного воздействия постоянным магнитным нолем с индукцией 400 мТ (2)
Продолжая перечисление возможных механизмов действия ЭМП на мозг, следует обратить внимание на межцентральные отношения. Наиболее чувствительным к ЭМП многие исследователи считают гипоталамус, где сосредоточены высшие вегетативные центры (рис. 17). Но имеется мнение, что гиппокамп может брать на себя ведущую роль в организации реакции мозга на ЭМП. Конечно, важную роль в осуществлении этой реакции играет кора больших полушарий мозга и другие его ведущие отделы.
Иерархия межцентральных отношений в мозге при действии ЭМП меняется в зависимости от исходного функционального соотношения отдельных центров, а для детального анализа этого соотношения необходимо знать, что происходит на клеточном уровне.
Не разбирая подробно роль нейрона и глии в реакции мозга на ЭМП (чему посвящена отдельная глава), мы должны основное внимание уделить мембранным процессам, функция которых окончательно не выяснена.
Выдвигаемые гипотезы о молекулярных механизмах функционирования биологических мембран должны удовлетворять принципу реципрокности протекания в мембране сопряженных молекулярных процессов, что обусловливает совершенно необычные с позиций классических представлений конкурентные взаимоотношения поглощения Са и фосфорилирования АДФ, переноса электронов и активации АТФазы в митохондриях, а в плазматических мембранах приводит к реципрокным соотношениям электрогенного натриевого насоса и аккумуляции К+, активности холино- и адренорецепторов.
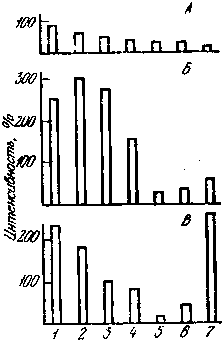
Рис. 17. Реакции разных отделов мозга кролика на ЭМП в норме (А), а также после введения кофеина (Б) или адреналина (В)
Ордината — интенсивность реакции (%); 1 — гипоталамус; 2 — сензомоторная кора; 3 — зрительная кора; 4 — специфические ядра таламуса; 5 — неспецифические ядра таламуса; 6 — гиппокамп; 7 — ретикулярная формация
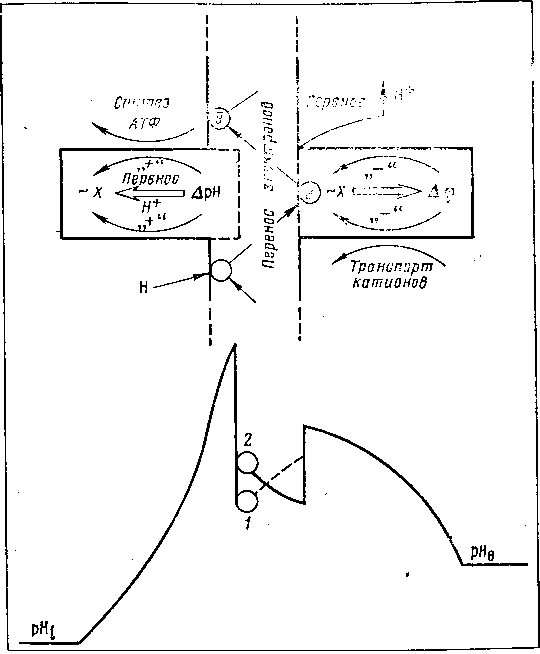
Этому требованию удовлетворяет гипотеза М. А. Шишло, согласно которой в мембране митохондрий происходит переход разности электрических потенциалов в скачок концентрации ионов водорода на границе раздела двух сред, который используется для синтеза АТФ (рис. 18). Система включает два основных элемента, работающих на основе электронно-колебательных взаимодействий, которые в соответствии с пространственно-зарядовой асимметрией мембраны, определяемой разностью pH по обе стороны и гидрофобными взаимодействиями белков и липидов, должны иметь положительную кооперативную связь ΔрН с виброном, участвующим в синтезе АТФ, и отрицательную кооперативную связь с вибронным возбуждением, осуществляющим выброс Н+ за пределы митохондрий и участвующим в транспорте кальция.
Из анализа модели вытекает важнейший вывод о том, что система, управляемая скачком концентрации протонов, не может быть системой с дипольными искажениями и должна обладать свойствами кристаллов, в которых параметром порядка служит деформация. Только в этом случае сброс протона по его электрохимическому градиенту может сопровождаться эффективной утилизацией энергии этого перехода в синтезе АТФ.
Если управление посредством изменения разности потенциалов вибронной неустойчивостью системы с дипольным искажением является общепризнанным и физически ясным процессом, то управление посредством ΔрН требует пояснения и доказательств.
Рис. 18. Схема механизма биологического действия ЭМП
Оно становится возможным лишь в том случае, если деформационная система стационарно находится в искаженном состоянии в результате химических гидрофобных взаимодействий. Быстрое появление скачка протонов на границе липид—белок нейтрализует гидрофобный градиент системы с деформационным искажением, что приводит к ее вибронному возбуждению, и протоны стекают по градиенту, участвуя в синтезе АТФ. Такой механизм обеспечивает суммацию в акте синтеза АТФ энергии деформационного искажения с энергией, выделяемой в ходе сбрасывания Н+ по их электрохимическому градиенту. Ни одна равновесная искаженная ян-теллеровская система без механизма поддержания искаженной конфигурации не может обеспечить такой суммации двух вкладов при активации эндергонической химической реакции синтеза АТФ. Из этого вытекает фундаментальное значение гидрофобных взаимодействий в мембранах.
Исходя из изложенного, ограничение процессов переокисления липидов становится особо необходимым для нормального функционирования АТФ-синтетазы и любого другого механизма, построенного на использовании энергии деформационного искажения кооперативных электронно-колебательных систем.
Становится очевидным, что гипероксия может избирательно повреждать функционирование этих деформационных кооперативных систем ослаблением гидрофобных взаимодействий при усилении перекисного окисления липидов.
В рассматриваемой системе изменение пространственно-зарядовой асимметрии мембраны изменяет вклад деформационной и электрической составляющих в элементарный акт синтеза АТФ, но не изменяет стехиометрию: один квант вибронного возбуждения системы с деформационным искажением — один элементарный акт синтеза АТФ. Изменяется при этом лишь форма и высота потенциальных барьеров для туннелирования протона и время протекания элементарного акта синтеза АТФ.
Интересно, что изменение трансмембранной ΔрН принципиально обращает пространственно-зарядово-временную симметрию системы с дипольными искажениями. Повышение pH закономерно приводит в такой системе к переключению от механизма поглощения К+ в обмен на Н+ к механизму электронного переноса К+, активируемого туннельным переносом электрона в данной структуре.
На этом мы закончим описание модели и покажем далее ее адекватность в описании эффектов ЭМП в митохондриях.
В рассматриваемой системе при воздействии ЭМП можно ожидать взаимодействия электрической составляющей с дипольной кооперативной системой мембран митохондрий. При этом ЭМП может инициировать ток деполяризации в сегнетоэлектрике, что должно активировать дыхательную цепь митохондрий и приводить в конечном итоге к диссипации поглощенной энергии в тепло. Система поддержания асимметрии мембраны за счет окислительно-восстановительного потенциала должна быть при этом активирована, и после выключения поля должен остаться «след» от компенсации деэнергизующего действия ЭМП.