У мохообразных с этим типом распространения во всех изолированных участках всегда встречается один и тот же вид. Это можно проиллюстрировать рядом примеров. Так, и в Европе, и на юге Сибири, и на юге Дальнего Востока произрастают Homalia trichomanoides, Leskeella nervosa, Mnium cuspidatum.
Еще один тип разорванного ареала. Вид встречается в Восточной Азии, включая юг советского Дальнего Востока, и потом, после очень большого перерыва, вдруг... появляется на Кавказе. Именно такое распространение имеет, например, один из представителей семейства Polytrichaceae — Pogonatum inflexum.
Нередко разрывы ареалов приходятся на моря и океаны. Это относится к очень многим видам из всех трех классов бриофитов, встречающихся в Приморье на Сахалине и на Курилах, а также на островах Японии. Таких видов, во всяком случае, несколько сот.
Но есть и более интересные формы разорванных ареалов, тоже приходящихся на воду. Так, значительная группа мхов и печеночников, распространенных в Восточной Азии (нередко включая также южную часть советского Дальнего Востока), после обширного перерыва появляются на западе Северной Америки. Иногда такое распространение называют амфипацифическим.
Очень часто разрывы ареалов имеют, так сказать, комбинированный характер: часть их приходится на сушу, часть на воду. Таких разрывов большинство. Один из примеров подобного разрыва — большая группа восточноазиатско-восточносевероамериканских видов. Они встречаются в Восточной Азии (очень многие — в пределах советской части Дальнего Востока) и в восточной (приатлантической) части Северной Америки, в восточных и юго-восточных штатах США. В западной половине Северной Америки, более близко расположенной к Восточной Азии, все эти виды отсутствуют. Цветковые растения с таким распространением на каждом материке представлены особыми видами. Общих видов почти нет. Мохообразные иногда тоже имеют разные (хотя и близкие) виды — на разных материках, но в большинстве случаев одни и те же, общие для Восточной Азии и восточной половины Северной Америки: Buxbaumia minakatae, Anomodon thraustus, Schwetschkeopsis fabronia, Aulacomnium heterostichum.
* * *
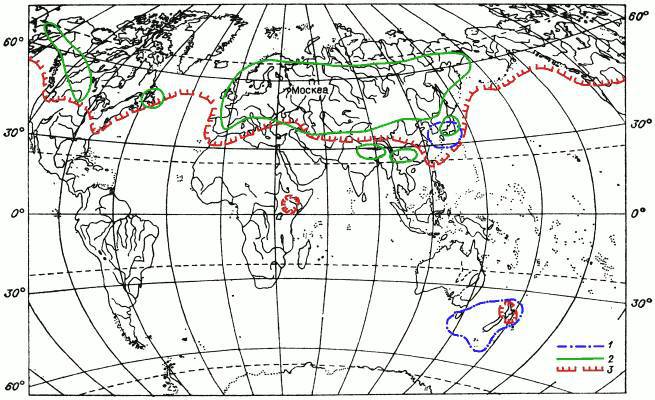
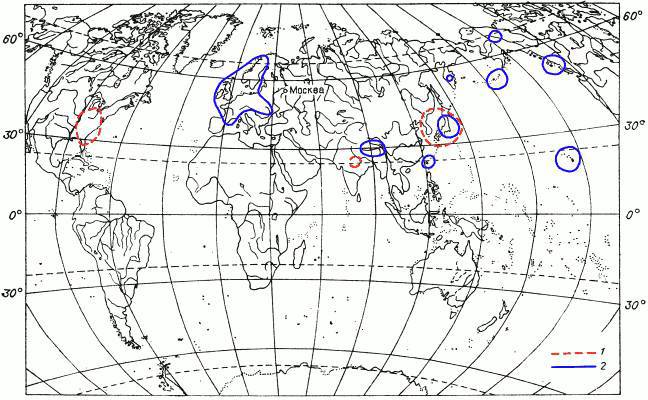
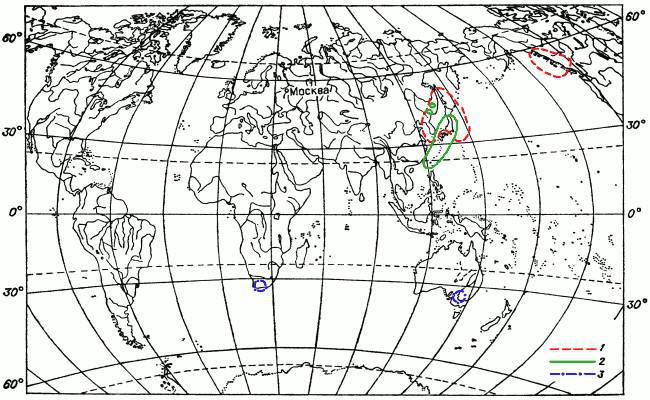
Вариантов дизъюнкций (разобщений) ареалов мохообразных множество. Перечислять их все мы не собираемся. Названы лишь некоторые примеры. Часть их проиллюстрирована картами ареалов (рис. 11—13), на которых можно увидеть и примеры других типов дизъюнкций, не упомянутых в тексте.
Рис. 11. Картосхема I распространения мохообразных.
1 — ареал печеночника Metzgeria furcata (пример дизъюнктивного ареала с дизъюнкцией в тропиках); 2 — ареал листостебельного мха Entodon concinnus (пример ареала широко распространенного в Северном полушарии вида); 3 — ареал листостебельного мха Hylocomium splendens (пример ареала широко распространенного в Северном полушарии вида, встречающегося в Южном полушарии).
Рис. 12. Картосхема II распространения мохообразных.
1 — ареал листостебельного мха Schwetschkeopsis fabronia (пример ареала восточноазиатско-восточносевероамериканского вида); 2 — ареал печеночника Anastrepta orcadensis (пример многократно разорванного ареала).
Рис. 13. Картосхема III распространения мохообразных.
1 — ареал листостебельного мха Pleuroziopsis ruthenica (пример ареала восточно-азиатско-западно-североамериканского вида); 2 — ареал листостебельного мха Hondaella. Монотипный род (пример ареала рода, имеющего ограниченное распространение); 3 — ареал печеночника Carrpos sphaerocarpos (пример ареала вида, распространение которого ограничено Южным полушарием).
Возникает вопрос, каким образом могли возникнуть подобные ареалы. Ведь исходным для любого ареала любого вида всегда и везде является монолитность, то есть ареал, состоящий только из одного куска. Все равно, большого или маленького. Откуда же берется несколько кусков?
В чисто теоретическом плане можно предположить, по меньшей мере, три способа, три пути создания дизъюнкций. 1. Виды в каждом изолированном участке возникают всякий раз заново, независимо друг от друга. 2. Изолированность создается дальним и сверхдальним разносом зачатков мохообразных (главным образом спор) с помощью воздушных течений. 3. Раздробленный на серию изолированных участков ареал представляет собою остаток некогда единого, сплошного (монолитного) ареала.
Рассмотрим каждый из этих возможных путей.
Первый путь. Многократное возникновение вида. Исходя из того, что мы сегодня знаем об эволюции и механизме видообразования, многократное возникновение вида можно представить лишь с очень большим трудом. И уж вовсе маловероятным кажется допущение о сколько-нибудь широких масштабах этого явления. Вообще этот путь — скорее логическое допущение, чем основанный на реалиях вывод. Короче говоря, этот путь создания разорванных ареалов мохообразных мы должны сразу же и решительно отбросить. Неприемлем он и в отношении других растений.
Второй путь. Дальний и сверхдальний разнос зачатков. Здесь надо рассмотреть по крайней мере два варианта — вид растет на суше, в глубине континента (первый вариант) и вид растет у берега моря (второй вариант). В обоих случаях разнос будет происходить преимущественно в направлении господствующих ветров (господствующих в сезон созревания спор или других зачатков). В отдельные моменты зачатки могут быть подняты на большую высоту, отнесены далеко и приземлиться на каком-либо участке, не контактирующем с границей ареала. На военном языке это называется десантом. Оставим пока в стороне вопрос о возможной приживаемости зачатков вдалеке от основного ареала. Важно отметить, что такой заброс зачатков на удаленный от местопроизрастания вида участок возможен.
Но если рассматривать разнос зачатков не как однократный акт, а как процесс, идущий непрерывно в течение всей жизни вида, а она исчисляется миллионами, десятками миллионов, а подчас, возможно, и сотнями миллионов лет, то станет совершенно очевидным, что такой разнос не может привести к формированию изолированных участков ареалов. Разнос зачатков, рассматриваемый во времени, в целом обратно пропорционален расстоянию. Чем дальше от растения, произведшего зачатки, находится участок, тем меньше на него попадет зачатков. И наоборот. Значит, наращивание, прирост ареалов должны происходить постепенно, шаг за шагом, без образования оторванных от основного участка фрагментов. Другими словами — сплошным фронтом.
Такой прирост будет происходить до естественной преграды. Преградой может служить, например, море. Здесь прирост ареала прекращается. А если там, «за синим морем», лежащим в направлении господствующих ветров, лежит остров или материк с условиями, более или менее соответствующими данному виду? Могут ли в этом случае диаспоры попасть туда, прижиться на новом месте и, таким образом, привести к возникновению разорванного ареала?
Чтобы ответить на этот вопрос, обратимся к результатам чрезвычайно интересных опытов голландского бриолога ван Зантена [Van Zanten, 1978], решившего проверить, существует ли связь между степенью устойчивости спор мхов к неблагоприятным воздействиям и характером географического распространения видов. Ученый исследовал споры ряда видов мхов, произрастающих в Южном полушарии, в отношении устойчивости к низким температурам и высушиванию. Именно устойчивость к названным факторам и определяет жизненность спор в условиях дальней и длительной транспортировки воздушными течениями. Споры одних и тех же видов подвергались суровым испытаниям в различных режимах: высушивались, часть их после этого выдерживалась в условиях низкой температуры (−30°C) в течение различного времени, часть увлажнялась. После этого споры проращивались. По проценту спор, не потерявших способности прорастать, можно было судить об устойчивости тех или иных видов.