А теперь зададимся вопросом: в какую сторону «читается» ген – к 3’ или 5’? Теперь, когда генные инженеры уверенно расшифровывают нуклеотидные последовательности и синтезируют их, это вопрос отнюдь не праздный.
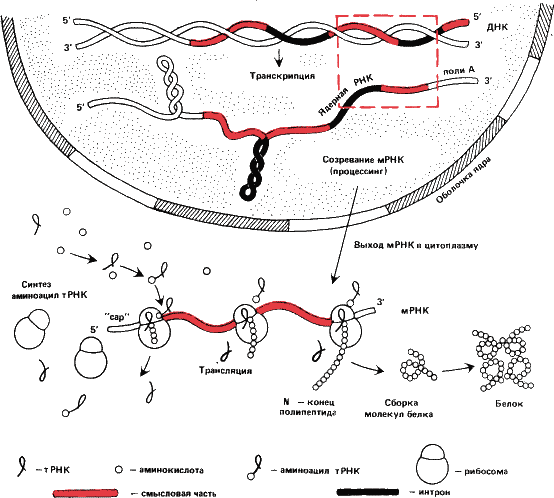
Рис. 17. Упрощенная схема передачи генетической информации с ДНК на белок. С находящейся в ядре ДНК снимается «рабочая копия» гена – гетерогенная ядерная РНК (этот процесс называется транскрипцией). Значительная, как правило, большая ее часть не кодирует аминокислотных последовательностей и отрезается ферментами – эндонуклеазами и отбрасывается. Тогда же вырезаются и «нечитаемые» вставки – интроны. Созревшая мРНК, получившая «шапочку» (cap – англ.) на 5’-конце и полиадениловую последовательность на З’-конце, через пору в ядерной оболочке выходит в цитоплазму (часто в виде комплексов с белком – информофер или информосом, на рисунке не показанных). В цитоплазме информация считывается с мРНК белоксинтезирующими аппаратами клетки – рибосомами (трансляция). Рибосомы гуськом идут по мРНК: каждый раз, когда рибосома смещается на три нуклеотида к З’-концу, к растущей полипептидной цепи прибавляется один аминокислотный остаток. Аминокислоты доставляются к рибосомам молекулами транспортной РНК (мРНК). Отдав аминокислоту, мРНК образует снова комплекс (аминоацил – мРНК) с другой и снова вовлекается в процесс. Полипептидная цепочка, оборвавшись на бессмысленном, терминирующем кодоне, свертывается специфичным образом. Это вторичная структура белка, поддерживаемая водородными связями; обычно это однонитчатая спираль (спираль Полинга – Кори). Спираль, в свою очередь, складывается, образуя третичную структуру. Наконец, многие белки состоят из более чем одной полипептидной цепи. Таков, например, гемоглобин, молекула которого состоит из четырех субъединиц. Это четвертичная структура.
Установлено, что полипептидные цепи в клетках синтезируются от N – конца к C – концу. Значит, у матричной РНК начало там, где кодируется N- конец. Оно соответствует 5’-концу РНК. В двойной спирали ДНК разобраться труднее. Дело в том, что слагающие ее нуклеотидные цепочки направлены в разные стороны, как говорят, антипараллельны. Иными словами, одна цепь направлена в сторону З’—5’, а другая 5’—З’.
Смысловая цепь, в которой закодирована аминокислотная последовательность «считывается» ферментом РНК-полимеразой с З’-конца. Образующаяся при этом мРНК, естественно, оказывается точным аналогом комплементарной цепи и будет начинаться с 5’-конца, с того, с которого начинается трансляция, то есть белковый синтез.
Но ведь с гена снимается не только «рабочий чертеж» мРНК. Ген и реплицируется, передаваясь из поколения в поколение, от матричной клетки к дочерним. Осуществляет этот процесс – репликацию – фермент ДНК полимераза.
Считается, что молекула ДНК-полимеразы садится на ДНК и движется по ней. При этом удваивается и смысловая цепь, и комплементарная к ней. Значит репликация смысловой цепи начинается с 3’-конца, как и транскрипция. Это аналогично тому как если бы мы перепечатывали текст с конца, а читали его, как и водится, с начала. В учебниках и популярных изданиях на это, как правило, не обращают внимания.
Последние годы ознаменовались сенсационными открытиями в изучении процессов репликации и трансляции. Природа подносила нам сюрприз каждый раз, когда начинало казаться, что уж теперь мы знаем об этих явлениях все.
Вот некоторые из сенсаций, за молодостью не попавшие в учебники.
Справедлива ли центральная догма? Мы уже упоминали, что генетическая информация передается от ДНК через РНК на белок, но не в обратную сторону. Это положение назвали центральной догмой молекулярной биологии. РНК-содержащие вирусы ее не нарушают. Просто у них выпадает начальное звено этой цели – ДНК. Генетическая информация передается из поколения в поколение закодированной в последовательностях РНК, с них же и считывается белок.
В принципе разница между ДНК и РНК не так уж и велика. Пентознофосфатный остов у РНК образует другой сахар – рибоза, который отличается от дезоксирибозы лишь наличием гидроксильной группы (OH). Набор оснований тот же, за тем исключением, что вместо тимина (5-метилурацила) в РНК содержится урацил (тот же тимин, только неметилированный). Недаром в природе встречаются ДНК, в состав которых входят и дезоксирибозы и рибозы. Такова, например, ДНК вируса герпеса, от которого на губах «высыпает лихорадка». Энергетические фабрики клеток – митохондрии – в значительной степени генетически автономны от ядра, они имеют свой геном, похожий на бактериальный. ДНК этого генома также содержит рибозу – от десяти до тридцати остатков на молекулу.
Все это не нарушало стройную догму. Тем большее смятение вызвало открытие синтеза ДНК на РНК. С. М. Гершензон писал еще в 1960 году о возможности подобного процесса, однако лишь сравнительно недавно был получен в значительных количествах фермент ревертаза (обратная транскриптаза), осуществляющий эту реакцию. Теперь этот фермент – обычный инструмент генных инженеров. Теперь мы можем дополнить центральную догму:
Например, РНК-содержащий вирус птичьего миэлобластоза может в результате обратной транскрипции стать ДНК-содержащим, встроиться в геном цыпленка и вызвать злокачественное перерождение клеток. Какую роль играет синтез ДНК на нити РНК в мире высших организмов, нам пока еще неизвестно.
Смысловая цепь: одна или две? Каких-нибудь пять лет назад все мы были твердо уверены, что матричная РНК синтезируется только на одной из двух цепей ДНК, получившей название смысловой. Вторая, комплементарная ей цепь нужна лишь для репликации ДНК и репарации – «починки» поврежденных участков. Если, например, жесткая радиация вырвет кусок из одной из цепей двойной спирали, специальные ферменты – репаразы заполняют брешь, пристроив на ее месте последовательность нуклеотидов, комплементарную оставшейся.
И тем не менее в последнее время появились данные, свидетельствующие о том, что в геноме дрозофилы, например, синтез РНК может идти на обеих цепях ДНК. Это так называемый симметричный синтез. Любопытно, что он идет от одной точки в разные стороны: в каждой цепи от З’ к 5’-концу, так что обе образующиеся РНК начинаются с 5’-конца. Биологический смысл этого явления мы еще не знаем.
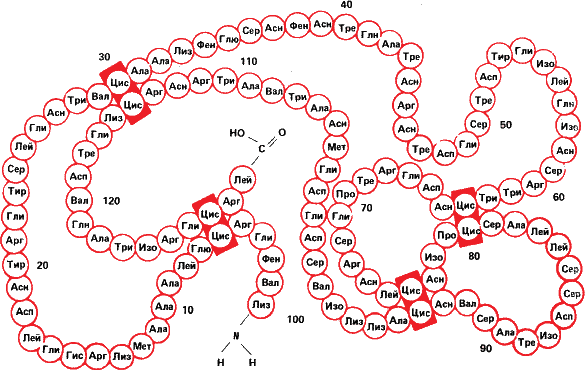
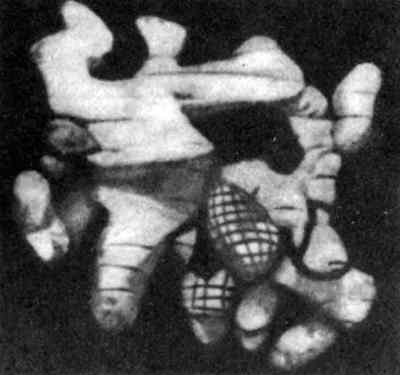
Рис. 18. Вверху – первичная структура белка лизоцима, разрушающего оболочки бактерий. Обратите внимание на четыре сшивки цис – цис (дисульфидные связи, которые мы уже видели на рис. 3). Внизу же не абстрактная скульптура, а модель третичной скульптуры лизоцима, полученная методом рассеивания рентгеновских лучей.
Перекрывается ли код? Первые гипотетические модели кода были перекрывающимися Это значило, что последовательность нуклеотидов могла кодировать разные аминокислотные последовательности, в зависимости от начала считывания. Так, последовательность АТТГЦАТЦГ, если считывалась бы с А, кодировала бы Тир—Вал—Ала, если со второго нуклеотида (Т)—Вал—Ала—Мет, и т. д. Подобный код накладывал бы жесткие ограничения на последовательность аминокислот в белках. И все облегченно вздохнули, когда С. Бреннер доказал, что каждый триплет нуклеотидов в ДНК и РНК считывается только один раз (неперекрывающийся код).
И опять как гром с ясного неба. Оказалось, что у некоторых фагов гены перекрываются. У фага φX174 имеется двойное перекрытие, у фага G4 даже тройное, то есть с одной нуклеотидной последовательности считываются три аминокислотных! Это предел экономичности сигнала. Фагам хорошо, а вот каково молекулярным генетикам? Как широко распространен этот феномен в мире вирусов? Встречается ли он у высших организмов? Не знаем.