Продвижение вперед идет очень маленькими, очень неуверенными шажками, временами возникающие неумеренные надежды сменяются горькими разочарованиями. Порой конструктивный, «рабочий» пессимизм перерастает в отчаяние: нет-нет кто-нибудь из конформаторов выскажется в том смысле, что взялись мы-де за принципиально неразрешимую задачу, что все это слишком сложно и для наших маломощных вычислительных машин, и для наших примитивных умов…
Позвольте, последнее ведь где-то уже было. Ну конечно, Г. Уэллс, «Машина времени». Путешественник во времени рассказывает о том, как его машина, позволяющая перемещаться в прошлое и будущее, попала в руки морлоков — отдаленных обезьяноподобных потомков человека, обитающих под землей. «Морлоки даже разбирали машину по частям, стараясь своим слабым разумом понять ее назначение».
Пытаясь своим слабым разумом понять назначение… Нет, нет, это только минутная депрессия. Как ни скромны итоги каждой очередной попытки, они все же хоть немножко прибавляют уверенности в конечном успехе, становятся еще одним кирпичиком, использованным при возведении здания теории третичной структуры. Общие контуры этого здания еще совершенно неясны, но о некоторых деталях можно кое-что уже сказать.
Начнем с самой элементарной проблемы: гибкости белковой цепи. Можно предположить, что в зависимости от того, какие именно аминокислотные остатки входят в те или иные ее участки, должна различаться также и гибкость этих участков. В принципе гибкость белковой молекулы обусловлена возможностью вращения отдельных ее частей вокруг некоторых валентных связей.
При таком вращении происходит взаимное сближение или удаление каких-то атомов, что и определяет их притяжение или отталкивание в зависимости от разделяющего их расстояния, сорта атомов, наличия электрического заряда, — словом, смотри предыдущую главу. В силу межатомного взаимодействия, и прежде всего именно в силу атом-атомного отталкивания, вращение вокруг отдельных связей оказывается более или менее «заторможенным»: существуют такие положения, при которых энергия отталкивания очень велика.
Поскольку же ближайшее атомное окружение связей, вокруг которых возможно вращение, может быть разным у различных аминокислотных остатков, естественно, неодинаковой оказывается также и степень «заторможенности».
Внутримолекулярные ситуации, при которых межатомное отталкивание велико, не могут реализоваться в действительности (почему именно, мы уже говорили) и называются стерически запрещенными. Это означает, что в результате вращения вокруг определенной связи две разделяемые ею части молекулы могут принимать друг относительно друга не все возможные положения, допускаемые таким вращением, а лишь некоторые из них, так называемые «стерически разрешенные». И конечно, можно ожидать, что наборы стерически разрешенных положений будут различаться от остатка к остатку.
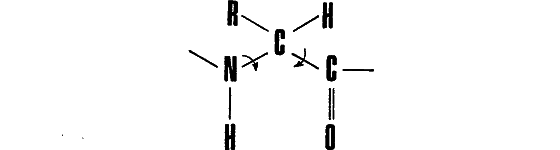
Если рассмотреть элементарное звено белковой цепи, соответствующее некоторому аминокислотному остатку (так называемую дипептидную единицу), окажется, что вращения в главной цепи возможны вокруг двух связей:
(Буквой R здесь обозначена боковая цепь аминокислотного остатка.)
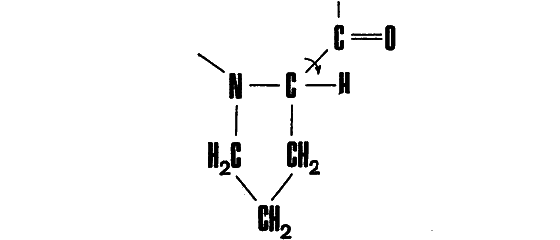
Сразу же можно указать на исключение из этого правила — остаток пролина:
в котором вращение вокруг связи N ― С невозможно: определенный угол поворота зафиксирован жестким кольцом. В месте включения этого остатка белковая цепь оказывается, таким образом, более жесткой.
Напротив, у остатка глицина доля стерически разрешенных конформаций высока, поскольку роль бокового радикала выполняет «маленький» атом водорода. И если пролин — наименее гибкий остаток, то глицин — наиболее гибкий из всех двадцати.
Все это, повторяем, можно сообразить сразу, безо всяких расчетов, лишь взглянув на химические формулы. А вот точное определение степени и, главное, характера гибкости каждого остатка, его возможности принимать определенные конформации, — это уже требует расчета. И именно описание «конформационной гибкости» отдельных остатков оказалось первой задачей теоретического конформационного анализа белков, задачей, которая была разрешена без особого труда, хоть и не обошлось в среде конформаторов без споров о том, какой из остатков более, а какой менее гибкий. Споры эти, между прочим, как две капли воды напоминали перебранку, подслушанную двумя героями И. Ильфа и Е. Петрова в месткоме некоего учреждения:
«— …Лыжная вылазка проведена недостаточно. А почему, товарищи? Потому что Зоя Идоловна проявила недостаточную гибкость.
— Как? Это я недостаточно гибкая? — завопила ужаленная в самое сердце Зоя.
— Да, вы недостаточно гибкая, товарищ!
— Почему же я, товарищ, недостаточно гибкая?
— А потому, что вы, товарищ, совершенно негибкая.
— Извините, я чересчур, товарищ, гибкая.
— Откуда же вы можете быть гибкая, товарищ?»
И еще мною часов спустя из дальней комнаты слышались голоса:
— Я, товарищ, чересчур гибкая!
— Какая же вы гибкая, товарищ?
До такого накала страстей среди конформаторов, правда, не доходило, но смысл некоторых дискуссий о расчетных оценках конформационной подвижности различных аминокислотных остатков, вне всякого сомнения, можно было бы передать примерно теми же выражениями:
— А я вам говорю, товарищ, что дипептидная единица аспарагина более гибкая, чем аланина!
— Откуда же она может быть гибкая, когда она совершенно негибкая, товарищ?
(Не следует считать пространное отступление исключительно порождением желчного нрава авторов: напротив, оно скорее относится к разряду лирических воспоминаний. Ведь конформационные расчеты дипептидных единиц и споры на эту тему отшумели, по меркам современной молекулярной биологии, очень давно, лет пять тому назад, и, стало быть, представляют собой незабвенное босоногое детство теоретического конформационного анализа пептидов и белков. А об этой идиллической поре, когда казалось, что значительная часть проблемы поиска третичной структуры белка сводится к расчету стабильных структур дипептидных единиц аминокислотных остатков, авторам всегда приятно вспомнить. Тем более что и сейчас нет-нет, да и появится в каком-нибудь научном журнале статья, рассказывающая об очередном варианте расчета какой-либо дипептидной единицы. Непосредственность и эмоциональность, сопровождающие зачастую подобные статьи, близки и понятны чутким сердцам авторов: никому, и даже самым суровым из конформаторов, не хочется расставаться с детством.)
Конформационные расчеты дипептидных единиц — первые робкие попытки подступиться к третичной структуре — принесли все же значительную пользу. На их основе удалось классифицировать основные типы поворотов, изгибов остова белковой цепи в месте включения отдельных аминокислотных остатков. Группа итальянских исследователей даже поспешила назвать такого рода классификацию стереохимическим кодом третичной структуры белка. Что же, стремление придумать еще один молекулярно-биологический код (именно код, это так хорошо звучит!) можно было бы только приветствовать, но использование именно этого термина в данной ситуации неуместно. Под кодированием, как мы помним, обычно понимают способ преобразования определенной информации, а здесь ни о чем подобном речи нет. Правильнее говорить о стереохимическом или конформационном алфавите, на котором может быть записана третичная структура, да и то с известной осторожностью: расчеты дипептидных единиц показывают, что такой алфавит описывает структуру лишь приближенно, с точностью до какого-то интервала «разрешенных» межатомными взаимодействиями значений углов внутреннего вращения.