Тут я снова слышу восклицание: «Стоп-стоп! А почему же вы тогда людей не включили в эту группу видов, неразборчивых в своих сексуальных связях?» Ответ: потому что я был достаточно осторожен, чтобы употребить слово «обычно». Да, большинство женщин на протяжении своей жизни имеет более одного сексуального партнера. Более того, многие женщины иногда имеют несколько партнеров одновременно. И тем не менее норма для женщины — в течение одного репродуктивного цикла вступать в сексуальные отношения только с одним мужчиной, тогда как норма для самки бонобо или верветки — спаривание с несколькими самцами.
ЭТАП 3. На следующей стадии исследования Силлен-Тульберг и Мёллер объединили данные, полученные на первом и втором этапах, чтобы выяснить — прослеживается ли какая-нибудь связь между степенью открытости овуляции и той или иной моделью брачного поведения? Если упрощенно воспринять описанные выше конкурирующие теории, то скрытая овуляция должна быть более характерна для моногамных видов (если, конечно, верна «теория папы-домоседа»), а демонстрируемая — для видов, практикующих промискуитет (беспорядочные сексуальные отношения). Выяснилось, что скрытая овуляция характерна для подавляющего большинства моногамных видов приматов, она отмечается у 10 видов из 11. Ни у одного из моногамных видов не бывает резко выраженных внешних признаков овуляции, в то же время из 18 видов с очевидной овуляцией для 14 характерно беспорядочное сексуальное поведение. Это, по-видимому, служит сильным аргументом в пользу «теории папы-домоседа».
Тем не менее этот аргумент лишь отчасти подтверждает теорию, потому что отсутствует обратная корреляция. Хотя у большинства моногамных видов отмечается скрытая овуляция — она, в свою очередь, вовсе не гарантия моногамии. Из 32 видов со скрытой овуляцией 22 не моногамны: для них характерен или промискуитет, или гаремы. К видам со скрытой овуляцией относятся моногамные дурукули (ночные обезьяны), преимущественно моногамные люди, живущие гаремами лангуры и образец промискуитета — верветки. Таким образом, какова бы ни была эволюционная причина скрытой овуляции, она может сочетаться с самыми разными брачными системами.
Точно так же, хотя большинство видов с демонстрируемой овуляцией ведут беспорядочную сексуальную жизнь, сама эта беспорядочность не является гарантией демонстрируемой овуляции. В самом деле, для большинства — у 20 из 34 — видов приматов, практикующих промискуитет, характерна овуляция либо скрытая, либо сопровождаемая слабо выраженными внешними признаками. Точно такая же картина наблюдается у гаремных видов: овуляция у них может протекать и совершенно незаметно, и малозаметно, и с явными внешними признаками. Такая сложность подсказывает нам, что скрытая овуляция может выполнять различные функции в различных брачных системах.
ЭТАП 4. Чтобы определить, как именно меняется эта функция, Силлен-Тульберг и Мёллер предложили здравую идею — надо изучить генеалогическое древо существующих видов приматов. Таким образом ученые надеялись определить те точки в эволюционной истории приматов, в которых происходили изменения в овуляционных сигналах и брачных системах. Исследователи исходили из того, что некоторые виды современных приматов, очень близкие друг другу, а значит, вероятно, лишь недавно разделившиеся, тем не менее практикуют различные системы брачных отношений и овуляционных сигналов. Это означает, что и эволюционные изменения в этих системах произошли сравнительно недавно.
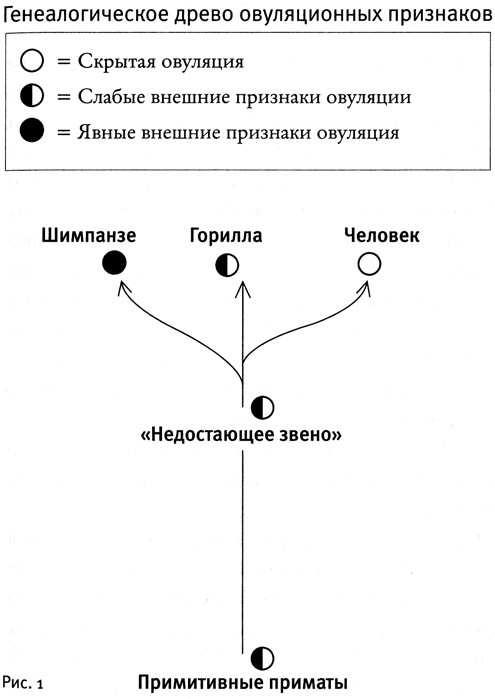
Вот пример того, как работает подобная аргументация. Известно, что люди, шимпанзе и гориллы генетически идентичны на 98 % и происходят от одного общего предка, так называемого «недостающего звена», жившего примерно девять миллионов лет назад. Тем не менее эти три современных потомка «недостающего звена» демонстрируют все три разных типа овуляционных проявлений: скрытую овуляцию у человека, слабо выраженные признаки у гориллы и сильно выраженные у шимпанзе. Отсюда можно сделать вывод, что лишь у одного из потомков «недостающего звена» система овуляционных сигналов такая же, как у предка, тогда как у двух других потомков должны были развиться отличающиеся системы.
Мы знаем, что у большинства видов примитивных приматов внешние признаки овуляции выражены слабо. Вероятно, таковы же они были и у «недостающего звена», а от него эта черта передалась горилле (см. рис. 1).
Однако в течение последних девяти миллионов лет у человека развилась скрытая овуляция, а у шимпанзе, напротив, овуляция с ясными внешними сигналами. Таким образом, эволюция овуляционных сигналов у человека и у шимпанзе пошла в противоположных направлениях от нашего общего предка со слабо выраженными внешними признаками овуляции. С нашей, человеческой точки зрения, набухшие «задние фасады» шимпанзе очень похожи на соответствующие части тела у павианов. Однако у шимпанзе и павианов эти признаки овуляции, должно быть, развивались совершенно независимо, поскольку предки павианов и предки «недостающего звена» разошлись примерно 30 миллионов лет тому назад.
По тому же самому принципу на генеалогическом древе приматов можно найти и другие узловые точки изменений в овуляционной системе. В ходе эволюции приматов случилось около двадцати таких изменений. Можно насчитать по крайней мере три независимых источника ярко выраженных овуляционных признаков (в том числе шимпанзе), по меньшей мере восемь независимых источников скрытой овуляции (включая нас, людей, орангутанов и шесть прочих групп обезьян), а также несколько случаев повторного возникновения слабо выраженных овуляционных сигналов, которые развились либо из скрытой овуляции (например, обезьяны ревуны), либо из ярко выраженной демонстрационной (многие макаки).
Подобно тому, как мы выделили узловые точки в процессе развития овуляционных сигналов, можно найти подобные точки и в эволюции брачных отношений. Общий предок всех высших приматов вел, скорее всего, беспорядочную половую жизнь. Однако если мы взглянем на брачные системы людей и наших ближайших родственников, шимпанзе и горилл, то мы увидим все три типа брачных отношений: гарем у горилл, промискуитет у шимпанзе, моногамия либо гарем у людей (см. рис. 2.).
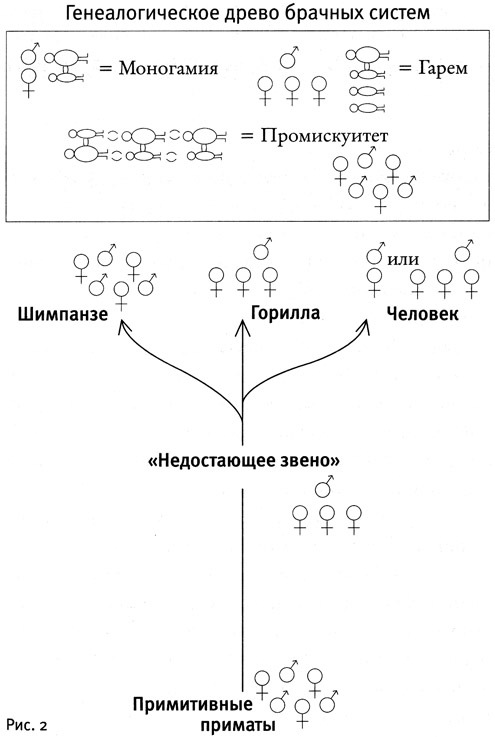
Таким образом, получается, что у двух из трех видов приматов, произошедших от одного общего предка («недостающего звена»), жившего девять миллионов лет назад, должно было измениться сексуальное поведение. Согласно некоторым данным, самки «недостающего звена» были объединены в гаремы, и, следовательно, гориллы и люди в некоторых сообществах просто унаследовали эту систему. Но шимпанзе в процессе дальнейшей эволюции вернулись к промискуитету, а у большинства людей, напротив, развилась моногамия. Мы видим, что и эволюция брачных систем у людей и шимпанзе шла в противоположных направлениях — как раньше мы это видели для овуляционных сигналов.
В целом похоже, что у высших приматов в ходе эволюции моногамия возникла в семи независимых случаях: у человека, у гиббонов и еще по крайней мере у пяти видов обезьян. Гарем сохранился в восьми случаях (считая и «недостающее звено»). Шимпанзе и по меньшей мере еще два вида обезьян вернулись к беспорядочным сексуальным отношениям уже после того, как их предки отказались от промискуитета в пользу гарема.
Итак, нам удалось воссоздать систему брачных отношений и тип внешних проявлений овуляции, которые существовали у приматов в далеком прошлом, и расположить все это на генеалогическом древе приматов. Теперь мы, наконец, можем объединить эти два массива информации и задать себе вопрос: какая же система брачных отношений преобладала всякий раз в той точке, где развивалась скрытая овуляция?