Рис. 4.1. Молекулы глюкозы и лактозы
Эти белки состоят из [галакзид — ]пермеазы, препровождающей молекулы лактозы через мембрану клетки, где они перевариваются; бета-галактозидазы, расщепляющей лактозу на глюкозу и галактозу; и [тиога-лактизид — ]трансацетилазы, чья роль еще полностью не выяснена.
Оперон ДНК — РНК — белки.
Представляется, что присутствие лактозы в клетке служит пусковым механизмом, приводящим в действие транскрипцию РНК, которая производит эти три белковых фермента. На самом же деле все обстоит значительно сложнее. Сигнал к производству различных белковых ферментов задается одновременно наличием лактозы и отсутствием глюкозы. Рассмотрим этот процесс на молекулярном уровне, чтобы выяснить его механизм.
ДНК порой представляют в виде обособленной молекулы, надежно защищенной благодаря своему крепкому сложению, хранящей жизненно важную для клетки информацию. Но это далеко не так. В действительности ДНК постоянно прощупывают, крутят, тормошат, раскрывают различные белковые ферменты. Такая деятельность заставляет эту информационную магистраль изрядно выкладываться.
Все эти действия обусловлены обликом ДНК и распределением электрического заряда. Двойная спираль имеет бороздки, маленькую и большую, а все нуклеотидные основания обладают только им присущим распределением электрического заряда (см.: Список идей, 6. Сборка модели ДНК, где рассказывается, как собирать часть ДНК из набора конструктора). Некоторые белки имеют размер и очертание, приходящиеся «впору» этим бороздкам. Благодаря распределению электрического заряда у белков и ДНК они могут плотно прилегать друг к другу. Однако притяжение не столь сильно, как ковалентные связи внутри каждой молекулы. Такое вкладывание одной молекулы в другую называют связыванием.
В зависимости от формы и распределения заряда белки присоединяются в соответствующих местах вдоль ДНК. Ввиду теплового движения молекул белки постоянно связываются и отделяются.
Соответствие сложных молекулярных очертаний часто представляется аналогичным ключу и замку. Лишь немногие очертания в достаточной степени соответствуют друг другу для соединения молекул. Белки тоже могут связываться с другими белками, образуя новую единицу под названием комплекс. Обычно комплекс приобретает иные по сравнению с исходной молекулой очертание и распределение заряда. Такую перемену, играющую главную роль в сборке белка, поскольку меняются «ключи» и «замки», именуют конформационным изменением.
РНК собирается с помощью белкового фермента (полимеразы), который прикрепляется к связывающей стороне ДНК, распускает двойную спираль посередине подобно «змейке» и переписывает (транскрибирует) порядок парных оснований ДНК на молекулу РНК. Затем РНК покидает ДНК и переносит (транслирует) порядок парных нуклеотидных оснований, собирая белок на молекулярном устройстве под названием рибосома. Каждая группа из трех нуклеотидных оснований, именуемая кодоном (см.: Список идей, 7. Кодоны), определяет, какую аминокислоту добавить к белку. Полимераза РНК связывается с ДНК лишь в тех местах, где приходится впору. Это прилаживание определяется не только очертанием молекулы полимеразы, но и наличием места связывания у ДНК, которое в свою очередь зависит от изгибов ДНК.
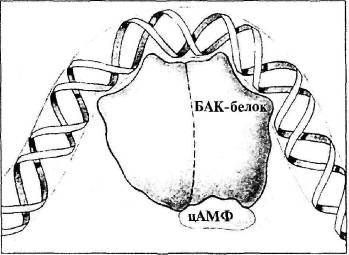
Для получения полной картины метаболического процесса на основе лактозы недостает трех молекул. Прежде всего, это белок-активатор катаболизма (БАК-белок). В обычном состоянии строение БАК-белка не позволяет ему соединяться с ДНК. БАК-белок содержит место для связывания с другой молекулой, циклическим аденозинмонофосфатом (цАМФ). Молекула цАМФ вырабатывается в среде, где отсутствует глюкоза. Если цАМФ связан с БАК-белком, БАК-белок претерпевает конформационное изменение, позволяющее ему присоединяться к ДНК. В свою очередь, связывание комплекса БАК-белок/цАМФ с ДНК Е. coli заставляет ДНК сгибаться, как показано на рис. 4.2.
На заключительном этапе требуется наличие другого белка, действующего в качестве репрессора. В данном случае он называется lac-репрессором. Эта молекула обычно входит в бороздку ДНК в том месте, где нужно помешать закрепиться полимеразе РНК, переписывающей информацию ДНК на белки, которые усваивают лактозу.
Если лактоза не соединена с lac-репрессором, репрессор точно входит в бороздку ДНК в нужном месте, препятствуя выполнению полимеразой РНК возложенной на нее задачи перезаписи (транскрипции). Если лактоза соединена с lac-репрессором, это вызовет в репрессоре конформационные изменения, так что он уже не будет подходить бороздке ДНК Е. coli и не будет мешать полимеразе ДНК выполнять транскрипцию.
Рис. 4.2. Связь цАМФ с БАК-белком, а БАК-белка — с ДНК (из кн.: Raven P. H., Johnson G. В. Biology)
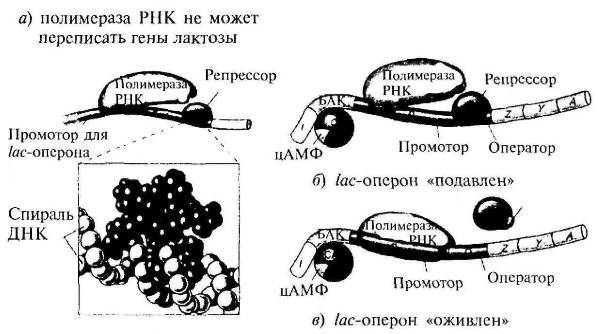
Рассмотрим, как эти молекулы сотрудничают, определяя наблюдаемую линию поведения Е. coli. В исходных условиях имеется глюкоза и отсутствует лактоза. При наличии глюкозы цАМФ не вырабатывается, а значит, не образовывается комплекс БАК-белок/цАМФ, не сгибается ДНК и полимераза РНК не переписывает белки для усвоения лактозы. Помимо этого, репрессор находится на ДНК, препятствуя соединению полимеразы РНК с ДНК в этом месте. Получается двойная блокировка перезаписи (рис. 4.3 а).
В смешанной среде с глюкозой и лактозой присутствие глюкозы препятствует образованию комплекса БАК — белок/ цАМФ, так что ДНК не изгибалась, а полимераза РНК не занималась перезаписью. Даже если присутствие лактозы вынудит репрессор покинуть бороздку ДНК, полного связывания полимеразы РНК не произойдет. Она покидает ДНК, так и не прикрепившись ни к одному из участков lac-оперона.
Рис. 4.3. Lac-peпpeccop (из кн.: Raven P. H., Johnson G. В. Biology)
В отсутствие глюкозы и лактозы появляется комплекс БАК-белок/цАМФ, изгибается ДНК в ожидании РНК, но при этом наличествует и репрессор. С точки зрения Е. coli отсутствие пищи означает предстоящий голод. Но мы видим, насколько она готова к возможному повороту событий. Если появится глюкоза, она не станет расходовать энергию на выработку белковых ферментов, а сразу приступит к усваиванию глюкозы. Если же появится лактоза, изогнувшаяся ДНК уже готова к сборке нужной РНК, стоит лишь лактозе соединиться с репрессором, который тотчас покидает ДНК (рис. 4.3 б).
В отсутствие глюкозы и при наличии лактозы происходит следующее. Недостаток глюкозы приводит к образованию комплекса БАК-белок/цАМФ, который соединяется с ДНК, вынуждая ее изгибаться. Это дает возможность полимеразе РНК отыскать свое место прикрепления. Присутствие лактозы приводит к ее связыванию с lac-репрессором и отсоединению репрессора от ДНК, так что уже вся полимераза РНК может присоединиться к ДНК и собрать три белка для усвоения лактозы (рис. 4.3 в).
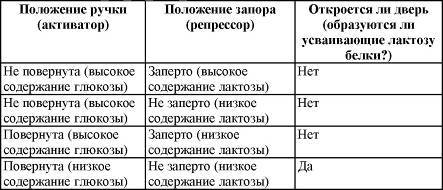
Подобное положение дел сходно с ситуацией с дверью, снабженной ручкой и засовом. Ручка действует подобно активатору, а запор выступает в роли репрессора. В таблице действие системы «ручка — засов» сравнивается с механизмом «активатор — репрессор».
Эта сложная система управления схожа со старым устройством Руба Голдберга, [8]где целая цепь сложных событий служит для достижения некой простой цели (рис. 4.4). И все же почему при всей затратности данного механизма Е. coli не вырабатывает постоянно все нужные ферменты, чтобы усваивать любой поступающий к ней сахар? Возможно, некогда такая бактерия и существовала.
Но появившаяся в ходе мутаций Е. coli с ее lac-опероном благодаря значительно большему по сравнению со своей старшей родственницей коэффициенту полезного действия вытеснила ее. Наглядный пример классического естественного отбора.
Белковые ферменты собираются практически одновременно с перезаписью РНК, когда РНК все еще прикреплена к длинной кольцевой молекуле ДНК. Поскольку Е. coli относится к прокариотным клеткам, у нее нет тормозящих ход метаболизма ядра или ядерной мембраны, так что усвоение лактозы начинается очень скоро. Е. coli прекрасно живет и на лактозе, и на глюкозе.