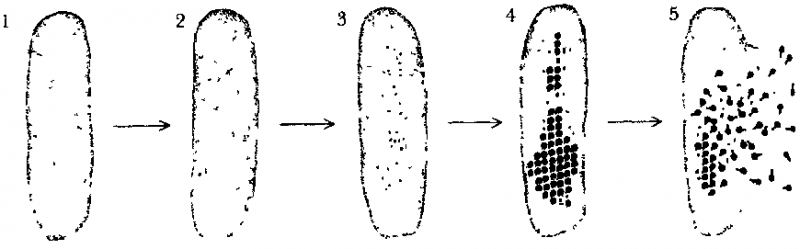
Время Происходящее действие
0 мин. Введение виральной ДНК.
1 мин. Порча хозяйской ДНК. Прекращение производства клеточных белков и начало производства чужих (Т4) белков. Одними из первых производятся белки, управляющие воспроизводством чужой (Т4) ДНК.
5 мин. Начинается производство ДНК вируса.
8 мин. Начало производства структурных белков, которые сформируют «тела» новых фагов.
13 мин. Произведена первая полная копия агрессора (Т4).
25 мин. Лизосома (тип белка) атакует стенку клетки-хозяина, бактерия лопается, освобождая «двухсотняшек».
Таким образом, через каких-нибудь двадцать четыре или двадцать пять минут после того, как фаг Т4 вторгается в клетку Е. coli, эта клетка оказывается полностью подчиненной и разрушается. Оттуда вырываются около двух сотен точных копий вируса-агрессора — «двухсотняшки» — готовые атаковать новые клетки и разрушать их так же, как и первую.
Хотя с точки зрения бактерии подобные вещи представляют собой серьезную опасность, мы, с нашей точки зрения, можем интерпретировать это как игру между двумя игроками: агрессор, или игрок «Т» (названный по имени класса фагов Т, куда входят Т2. Т4 и другие), и игрок «К» (клетка). Игрок Т старается проникнуть в клетку и овладеть ею изнутри с целью самовоспроизводства. Игрок К старается защитить себя и уничтожить агрессора. Описанная таким образом, молекулярная игра Т-К напоминает макроскопическую игру Т-К, описанную в предыдущем Диалоге. (Читатель, разумеется, без труда поймет, какой игрок — Т или К — соответствует Черепахе Тортилле, а какой — Крабу.)
Рис. 102. Вирусная инфекция начинается, когда ДНК вируса попадает в бактерию ДНК, бактерии при этом портится, зато ДНК вируса начинает размножаться. Синтез составляющих вирус белков и поглощение их вирусом происходит до тех пор, пока клетка не лопается, освобождая частицы (Hanawault & Haynes «The Chemical Basis of Life», cтp. 230)
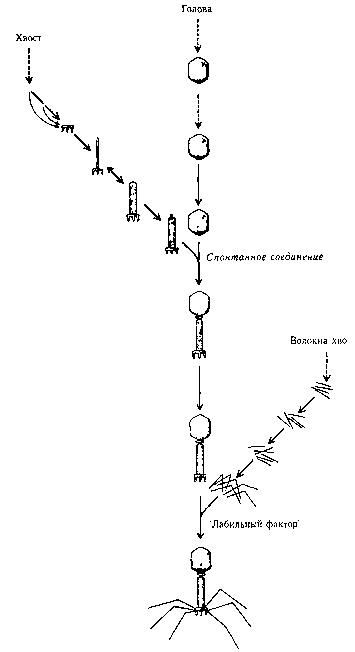
Рис. 103. В морфогенетическом пути вируса Т4 есть три основных ветви, ведущие к независимому образованию голов, хвостов и волокон хвоста, которые затем соединяются и формируют полные копии вируса (Hanawault & Hayness «The Chemical Basis of Life», cтp. 237)
Узнавание, маскировка и наклеивание ярлыков
Эта «игра» делает очевидным тот факт, что узнавание — одна из центральных тем клеточной и субклеточной биологии. Каким образом молекулы (или структуры высшего уровня) узнают друг друга? Чтобы энзимы работали хорошо, они должны быть способны присоединяться к определенным местам соответствующих субстратов; бактерия должна уметь отличать собственную ДНК от ДНК фагов; клетки должны узнавать друг друга и взаимодействовать определенным образом. Эта проблема узнавания может напомнить вам об основном вопросе формальных систем: как можно узнать, является ли данная строчка теоремой? Есть ли для этого разрешающая процедура? Подобные вопросы принадлежат не только области математической логики; они важны также в теории вычислительной техники и, как мы видели, в молекулярной биологии.
Техника ярлыков, описанная в Диалоге, является, на самом деле, одним из трюков, используемых Е. coli, чтобы перехитрить агрессоров-фагов. Идея заключается в том, что цепочка ДНК может быть химически отмечена путем присоединения к нескольким нуклеотидам маленькой молекулы — метила. Эта операция «наклейки ярлыка» не меняет основных биологических свойств ДНК, другими словами, метилированная (отмеченная ярлыком) ДНК может быть транскрибирована точно так же, как и неметилированная (не отмеченная ярлыком) кислота, таким образом, она может управлять синтезом тех же белков. Однако, если клетка-хозяйка обладает специальным механизмом, проверяющим, отмечена ли ДНК, то ярлык становится крайне важен. В частности, клетка может располагать системой энзимов, распознающих и уничтожающих неотмеченные цепочки ДНК. Найдя такую цепочку, эти энзимы безжалостно рубят ее на куски. В таком случае, увы всем непрошенным гостям!
Метиловые ярлыки на нуклеотидах можно сравнить со специальным типографским шрифтом. Используя эту метафору, можно сказать, что клетка E coli ищет цепочки ДНК, напечатанные этим «специальным шрифтом» и разрушает любую цепочку ДНК напечатанную иным «шрифтом». Контрстратегией фагов, разумеется, было бы научиться снабжать свою ДНК такими же ярлыками и, таким образом, заставить клетки в которые они вторгаются, воспроизвести эту ДНК.
Эта битва Т-К может продолжаться до произвольных уровней сложности, но мы не будем рассматривать ее дальше. Главное здесь в том, что это битва между хозяином, пытающимся не впустить ни одной чужой ДНК, и фагом, который старается ввести свою ДНК в какую-нибудь клетку, которая транскрибировала бы ее в мРНК (после чего ее воспроизводство было бы гарантировано). Можно сказать, что ДНК, которой удается таким образом воспроизвести себя, интерпретируется на высшем уровне так: «Меня можно воспроизвести в клетках типа X» (В отличие от упомянутого ранее бесполезного с точки зрения эволюции фага, в котором закодированы белки, его же разрушающие, подобный фаг интерпретируется «Меня нельзя воспроизвести в клетках типа X»
Суждения Хенкина и вирусы
Эти противоположные типы автореференции в молекулярной биологии имеют свою параллель в математической логике. Мы уже обсуждали математическую аналогию фагов-самоубийц — я имею в виду строчки Геделева типа, утверждающие собственную невозможность внутри определенных формальных систем. Однако, возможна и параллель с настоящим фагом, утверждающим собственную воспроизводимость в определенной клетке — суждение, утверждающее собственную воспроизводимость в определенной формальной системе. Суждения подобного типа называются суждениями Хенкина, по имени математического логика Леона Хенкина. Они строятся примерно так же, как Геделевы суждения — единственная разница заключается в отсутствии отрицания Мы начинаем, разумеется, с «дяди»:
Eа:Eа' <ПАРА-ДОКАЗАТЕЛЬСТВА-ТТЧ{а,а'}Λ ARITHMOQUINE{а'',а'}
и затем проделываем стандартный трюк. Предположим, что Геделев номер приведенного выше «дяди» — h. Арифмоквайнируя дядю, мы получаем суждение Хенкина:
Eа:Eа' <ПАРА-ДОКАЗАТЕЛЬСТВА-ТТЧ{а,а'}Λ
ARITHMOQUINE{SSS…SSS/a'',a'}>
. |____|
S повторяется h раз
(Кстати, видите ли вы, в чем это суждение отличается от —G?) Я привожу его целиком, чтобы показать, что оно не дает «рецепта» собственной деривации; оно просто утверждает, что такая деривация существует. Вы можете спросить, верно ли это утверждение? Существуют ли деривации суждений Хенкина? Действительно ли эти суждения являются теоремами? Полезно вспомнить, что не обязательно верить политику, провозглашающему: «Я честный», — это может оказаться как правдой, так и враньем. Достойны ли суждения Хенкина большего доверия, чем политики?
Оказывается, что суждения Хенкина всегда истинны. Хотя пока нам не совсем понятно, почему это так, придется нам здесь принять этот интересный факт на веру.
Явные и неявные суждения Хенкина
Я упомянул о том, что суждения Хенкина ничего не говорят о собственной деривации; они лишь утверждают, что такая деривация существует. Возможно придумать вариацию на тему суждений Хенкина — а именно, суждения, явно описывающие собственную деривацию. Интерпретацией на высшем уровне подобного суждения было бы не «существует некая последовательность строчек, являющаяся моей деривацией», а «описанная ниже последовательность строчек … является моей деривацией». Давайте назовем первый тип суждений неявным суждением Хенкина. Новое суждение, соответственно, будет явным суждением Хенкина, поскольку в нем содержится явное описание собственной деривации. Заметьте, что в отличие от своих неявных собратьев, явные суждения Хенкина не обязательно должны являться теоремами. На самом деле, очень легко написать строчку, которая утверждает, что ее деривация состоит из единственной строчки 0=0, — ложное утверждение, поскольку 0=0 не является деривацией чего бы то ни было. Однако возможно также написать явное суждение Хенкина, являющееся теоремой, — то есть суждение, в действительности дающее рецепт собственной деривации.