Во-первых, детальные палеоклиматологические[298] исследования показывают, что во время Ледникового периода в Центральной Азии существовали обширные пространства степной тундры, способные обеспечить всем необходимым охотников. Это подтверждается археологическими находками, отражающими основные виды занятий человека во время максимальной фазы последнего Ледникового периода (см. главу 6). Дэйл Гатри решительно выступает против гипотезы о полном отсутствии населения на юге, в сердце Мамонтовой степи.
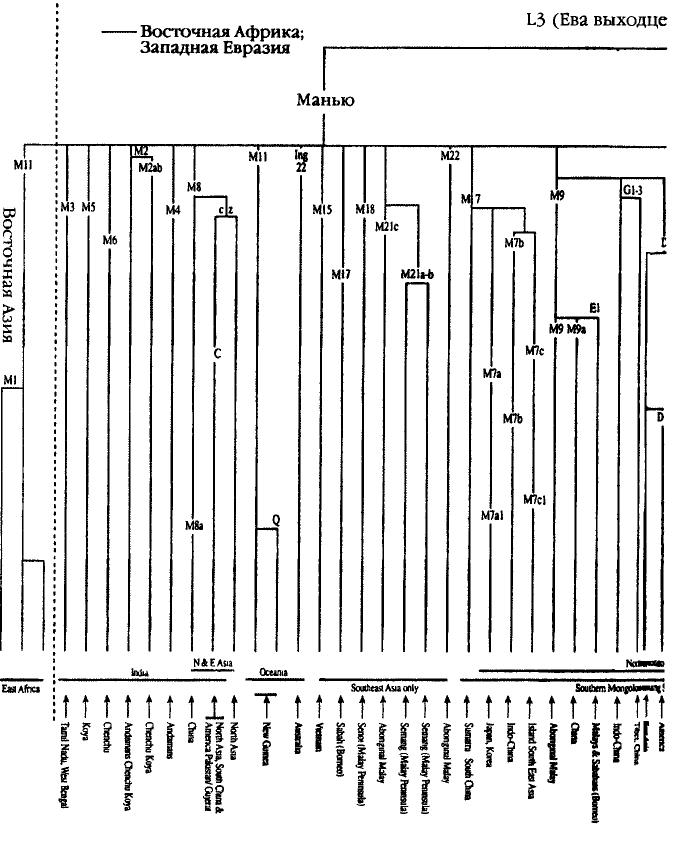
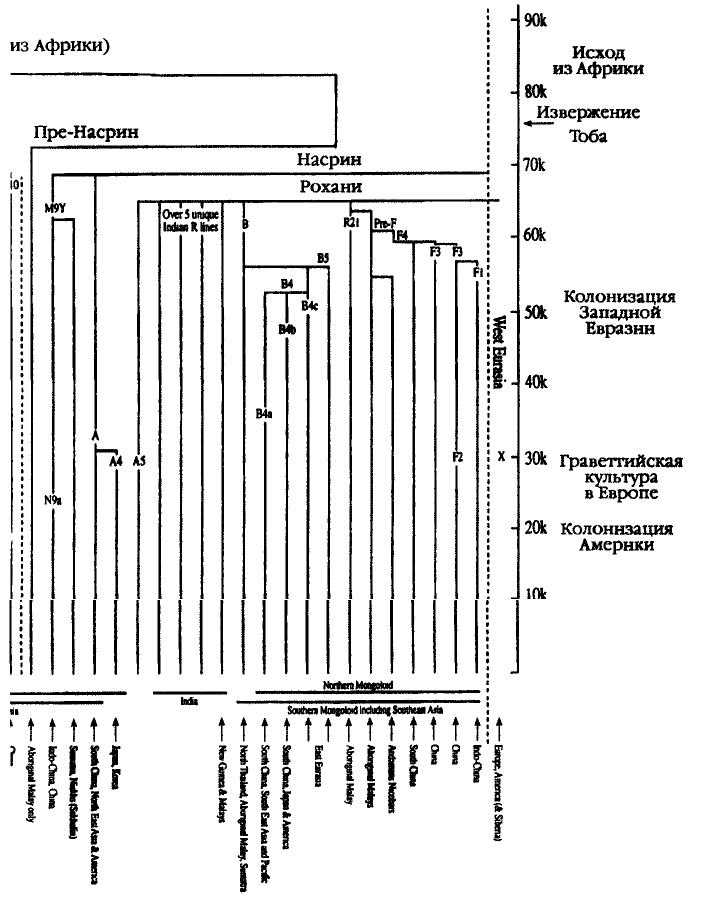
Во-вторых, как мы увидим ниже, подобная картина не позволяет объяснить наличие генетического многообразия и поливариантности на юго-западе Центральной Азии (в Узбекистане и Киргизстане) или резко выраженных географических различий между представителями разных ветвей генетического древа в Северной и Южной Азии. В частности, существует целая группа генетических линий, единых для Северной Европы, Северной Азии и Северной Америки, однако не имеющих ничего общего с Южным Китаем или Юго-Восточной Азией[299]. Схема развития генетического древа Как и в случае с историей исхода из Африки, ответ на интересующий нас вопрос станет более ясным, если мы посмотрим на маркеры Адама и Евы. Большим достоинством построения древа митохондриевой ДНК и Y-хромосомы является то, что они позволяют проследить ветви и даже побеги, соответствующие отдельным молекулам, применительно к каждому конкретному району. Эту увлекательную научную «игру» в гены и районы их распространения можно назвать филогеографией. В принципе она заключается в соединении разрозненных региональных побегов в крупные ветви и размещении последних на генетическом древе. Хотя правила предельно просты, их применение может оказаться достаточно сложным. Чтобы проследить векторы миграции из одного региона в другой, нам необходимо найти ветви-источники в обоих регионах и новый «побег» в искомом районе, который отсутствует на ветви прародины. Для прародины с многообразными векторами миграции в самых разных направлениях нам необходимо выявить общие ветви, имеющие разные «побеги» в других районах. Пожалуй, один из наиболее удачных примеров подобной методики был описан в работе видного итальянского генетика Антонио Торрони и его коллег, опубликованной в 1993 г. Правая часть Левая часть Рис 5.5 Эти исследователи выявили четыре основные линии митохондриевой ДНК этнической группы-родоначальницы коренного населения Америки, пометив их буквами А, В, С и D. Истоки каждой из этих буйно разросшихся ветвей, давших обильные «побеги» в Америке, можно возвести к аналогичным ветвям-прародителям А, В, С и D в Азии (см. главы 6 и 7). Мы можем применить этот же подход и к гораздо более сложной проблеме — идентификации прародины монголоидов в Азии. Взяв за начало митохондриевую ДНК Восточной Азии, мы можем заметить, что на сегодня там присутствуют девять четко сформировавшихся ветвей (гаплогрупп; см. рис. 5.5), которые можно обозначить буквами A, D, С, D, E, F, G, X и Z. За исключением сестринских групп С и Z, ни одна из этих групп не может быть выведена на основе другой. Каждая из них более или менее уверенно выстраивается на основе двух евразийских линий-родоначальниц — Манью или Насрин. Недавно были описаны еще несколько второстепенных групп (М7/М10 и N9), однако их существование никак не сказывается на значимости аргументации. (Более подробно о филогенезе и источниках по распределению гаплогрупп митохондриевой ДНК в Восточной Азии см. примечание[300].) Из девяти наиболее хорошо изученных восточно-азиатских ветвей митохондриевой ДНК три (С, X и Z, см. рис. 5.9) прослеживаются только к северу от условной линии «запад — восток», проведенной ниже Гималаев, тогда как остальные шесть (А, В, D, E, F и G) были обнаружены у обеих — как северной, так и южной — ветвей монголоидов. На первый взгляд может показаться, что эти уникальные северные ветви — весомые аргументы в пользу гипотезы о том, что прародина монголоидов находилась к северу от Гималаев, но на самом деле у этого феномена есть более простое объяснение. Дело в том, что линия С и близкородственная ей линия Z, а также загадочная группа X не являются коренными фигурантами древнейшей истории Монголии. Они появились среди северной ветви монголоидов откуда-то извне, вероятнее всего — из Сибири или Центральной Азии. Как мы скоро увидим, в поддержку этой версии существует целый ряд аргументов. Распределение генов и структура шести остальных ветвей генетического древа свидетельствуют о том, что генетическая прародина монголоидов находилась к югу от Гималаев. С особенной ясностью на это указывают древнейшие генетические ветви В и F. Таким образом, мы, по всей видимости, имеем дело сразу с несколькими источниками генов у жителей Юго-Восточной Азии, которые впоследствии распространились на север. Наоборот, среди обитателей Северной Азии мы имеем несколько ветвей, никак не связанных с коренным населением Юго-Восточной Азии. Возвращаясь к той же версии о развитии ветвей, можно отметить, что идея о южной прародине и последующей миграции монголоидов на север подтверждается данными исследования материнских генов и вполне соответствует гипотезе об эволюции сундадонтов в синодонтов (см. главу 5). Чтобы сосредоточить внимание на поисках возможного вторичного северного региона «дальнейшего развития той же темы», нам придется обратиться к обзору географических зон, где присутствуют все шесть ветвей митохондриевой ДНК — А, В, D, E, F и G, — являющихся общими для северной и южной ветвей монголоидов. Таким образом, продвигаясь с запада на восток, мы обнаружим, что в Центральной Азии присутствуют пять основных линий (за исключением G), в Тибете — все шесть, в Монголии — Пять (за исключением Е), в Китае — четыре (за исключением Е и G) и, наконец, в Корее — пять (за исключением[301]). Пожалуй, вряд ли можно считать простым совпадением, что страны с большим числом генетических линий, обычно ассоциируемых с монголоидной расой — Центральная Азия, Тибет, Монголия и Корея, — расположены вдоль 40-й параллели, которая, согласно Гатри, служила своего рода осью Мамонтовой степи (см. рис. 5.6). Основываясь на этих факторах, мы можем задать вопрос о том, каков примерный возраст этих ветвей, расположенных вдоль 40-й параллели. Тоомас Кивисилд установил, что местный возраст четырех из этих ветвей в Монголии весьма близок: ветвь А насчитывает 35 500 лет, В — 40 500 лет или 33 500 лет (см. ниже), D — 44 500 лет и, наконец, F — 42 500 лет. Эти даты значительно «моложе», чем средний возраст тех же линий на юге, в Южном Китае. Но самое важное заключается в том, что они вполне сопоставимы с датами начала активной колонизации Центральной Азии и относятся к эпохе после улучшения климата в Азии, которое произошло примерно 52 тысячи лет назад (см. главу 3) и благодаря которому первые обитатели Азии смогли проникнуть на просторы Мамонтовой степи[302].
И все же: по каким путям и маршрутам люди современного типа проникли в этот обширный регион? Обратившись к анализу двух древнейших линий Юго-Восточной Азии — В и F (идущих от потомков Насрин), которые тянутся из Юго-Восточной Азии в Тибет, Монголию и далее на северо-восток, вдоль побережья Тихого океана, мы видим, что они проникают в глубь материка в Сибири (Северная Азия), а затем останавливаются[303]. В этом отношении распределение этнических групп В и F отражает древнейшее распространение признаков сундадонтности. Как В, так и F достигают наиболее выраженной поливариантности среди туземных этнических групп Таиланда, Вьетнама и Камбоджи, что свидетельствует о том, что их прародиной является Индокитай. вернуться Палеоклиматология — научная дисциплина, изучающая состояние климата и прочих метеорологических факторов в давние эпохи истории Земли — Прим. перев. вернуться Палеоклиматологические исследования: см. карты на сайте http://www.esd.ornl.gov/projects/qen/NEW...MAPS/eurasia1.gif и /www.esd.ornl.gov/projects/qen/eurasia2.gif. Наличие генетического многообразия и поливариантности на юго-западе Центральной Азии: см. илл. 2 в Metspalu, E. et al. (1999) «Закавказье и распространение специфически кавказоидного типа митохондриевой ДНК» в кн. S.S.Papiha et al. «Многообразие геномов и его применение в генетике человека» (Kluwer Academic/Plenum Publishers, New York) cc. 121 — 134. Специальные карты на вебсайте Джонатана Адамса ESD ORNL /www.esd.ornl.gov/projects/qen/euras18k.gif и /www.esd.ornl.gov/ projects/qen/euras(2.gif. См. также археологические свидетельства, представленные в следующей главе. вернуться Группы А и X представляют собой первое поколение потомков N/Насрин, тогда как В и F происходят от Насрин через Рохани. Группы С, D, E, G и Z являются прямыми потомками М/Манью (см. илл. 5.9 и 5.10). Первое полное описание групп A-G, показанных на одном древе, было представлено в Torroni, A. et al. (1994) «Анализ митохондриевой ДНК на Тибете: его использование для установления происхождения тибетцев и их способности адаптироваться к условиям высокогорья» American Journal of Physical Anthropology 93: 189—99. О первоначальном наименовании группы X см. Forster, P. et al. (1996) «Происхождение и эволюция вариаций мтДНК у коренных народов Америки: пересмотр традиционной оценки» American Journal of Human Genetics 59: 935—45. Другая группа, Y, встречается почти исключительно на севере, у народности нивхов на о. Сахалин; эта группа очень молода и поэтому находится на периферии дискуссии (см. ниже). О группах Y и Z, а также о переоценке структуры древа азиатской мтДНК см. Kivisild, Т. et al. (2002) «Новые ветви и отрасли на древе мтДНК Восточной Азии» Molecular Biology and Evolution 19(10): 1737— 51; Yong-Gang Yao et al. (2003), указ. изд.; и илл. 2 в Forster, P. et al. (2003) «Эволюция мтДНК у аборигенов Азии и Папуа» в P.Bellwood and C.Renfrew (eds) «Исследования гипотезы о распространении сельского хозяйства и языка» (McDonald Institute for Archaeological Research, Cambridge), cc. 89—98. Главная задача заключалась в том, чтобы выявить различия между северной и южной генетическими линиями у монголоидного населения. С и Z — типы, связанные с группой М (о недавно выявленной «родительской» группе М8 см. Yong-Gang Yao et al. (2003), указ. изд.), и встречаются они почти исключительно в Северной Азии (а С также в Северной и Южной Америке), а не у южных монголоидов. Тип А, хотя он и распространен на северо-востоке Китая, в северо-восточной Сибири и Северной Америке (Kolman, C.J. et al. (1996) «Анализ митохондриевой ДНК у монголоидного населения и его использование для установления корней происхождения групп-родоначальников Нового Света» Genetics 142: 1321—34), отличается весьма частой встречаемостью и многообразием в южном и юго-западном Китае, на Тибете и Синьцзяне (см. Yong-Gang Yao et al. (2003), указ. изд.), но совершенно отсутствует в Индии и Западной Евразии (см. Kivisild, Т. et al. (1999) «Место индийских вариантов митохондриевой ДНК в глобальной системе материнских генетических линий и заселении Старого Света» в кн. S.S.Papiha et al. «Многообразие геномов и его применение в генетике человека» (Kluwer Academic/Plenum, New York) cc. 135—52. Таким образом, этнический источник в юго-западном Китае, с последующим продвижением по Янцзы на Тибет, представляется гораздо более вероятной прародиной А, чем прочие источники в Индии или Северной Азии. Группа Y, дочь N, выявлена только в Северо-Восточной Азии (Stariovskaya, Y.B. et al. (1998) «Многообразие мтДНК у чукчей Сибири: возможности применения для изучения генетической истории Древней Берингии и заселения Нового Света» American Journal of Human Genetics 63: 1473—91); Torroni, A. et al. (1993) «Вариации мтДНК у коренных жителей Сибири свидетельствуют о несомненной генетической близости с туземным населением Америки» American Journal of Human Genetics 53: 591—608). Однако возраст группы Y — всего лишь около 5000 лет, и она происходит от южноазиатской ветви N — N9. Распространение X, еще одной «дочери» N, ограничено европейцами и американцами (Brown, M. et al. (1998) «Гаплогруппа X мтДНК: древнее связующее звено между Европой/Западной Азией и Северной Америкой?» American Journal of Human Genetics 63; 1852— 61; кроме того, есть единичные сообщения о выявлении X у аборигенов Сибири (см. ниже). Хотя любая из этих трех линий (С, X и Z) теоретически могла стать характерной чертой генетических линий монголоидов, все эти линии полностью отсутствуют у южных монголоидов, но зато встречаются у других этнических групп, что делает их присутствие еще менее вероятным. В последнее время к этим группам присоединилось еще несколько кладов, выявленных в Китае: три из них происходят от М (М7, М9 и М10), а четвертый — от N (N9, примерный возраст 64300 ... 20 тысячи лет; является наследственным для Y): см. Yong-Gang Yao et al., указ. изд. Две азиатские «внучки» N (В и F) — единственные известные науке восточноазиатские потомки линии R/Poхани (не считая вновь описанного древнего клада, R9, возраст которого — 81 000 ... 24 600 лет; см. Yong-Gang Yao et al., указ. изд., и который является наследственным для F на юге и поэтому получил название F4 в Hill, С. et al. «Вариации митохондриевой ДНК у племени оранг асли на Малайском полуострове» American Journal of Human Genetics (готовится к печати); см. илл. 5.5). В и F отличаются наибольшим разнообразием и наивысшей встречаемостью у южных монголоидов Юго-Восточной Азии (Torroni, A. et al., указ. изд., Yong-Gang Yao et al., указ. изд.). Они выявлены также к северу от Гималаев в Центральной Азии (илл. 2 в Metspalu et al., указ. изд.), в Монголии (Kolman et al., указ. изд.), а также на Тибете, в Корее и Японии (Torroni, A. et al. (1994), указ. изд.; Horai, S. et al. (1996) «Полиморфизм мтДНК у населения Восточной Азии» American Journal of Human Genetics 59: 579—90). В то же время эти линии не встречаются на Крайнем Севере Сибири и в субарктическом регионе. Из этих южных линий В также выявлена в Америке, хотя опять-таки не в субарктическом регионе (см. главу 7). Так как линии В и F присутствуют по обе стороны Гималайского хребта, они являются потенциальными претендентами на роль линий-родоначальниц монголоидов, однако вопрос о том, присущи ли они всем монголоидам (линия F в Америке отсутствует), остается открытым. Мы рассмотрели три северноазиатские линии и три линии из Южной Азии, так что у нас остаются линии D, Е и G. Эти «дочери» М/Манью сохраняют потенциальную связь со своими истоками (дело в том, что они все восходят к М и имеют общую мутацию в быстрой (нестабильной) точке 16362 — см. Kivisild et al. (1999, 2002), указ. изд.). Линии D и G широко распространены на всей территории расселения монголоидов, а также в Индокитае. Уникальные локальные варианты D в большом количестве встречаются в Китае, Японии, Монголии, на Тибете, в Корее, Центральной и Северо-Восточной Азии, а также в Северной и Южной Америке (Torroni et al., (1994), указ. изд.; Torroni, A. et al. (1993) «Черты сходства с азиатами и континентальное распространение четырех ветвей-родоначальниц мтДНК коренных жителей Америки» American Journal of Human Genetics 53: 563—90; см. Kolman et al., указ. изд.; Kivisild et al. (1999, 2002), указ. изд. Хотя линия D и встречается в Южном Китае, она не характерна для Юго-Восточной Азии (Torroni et al. (1994), указ. изд.). Уникальные варианты линии G выявлены в Центральной Азии (см. илл. 2 в Metspalu et al., указ. изд.), на Тибете (Torroni et al., (1994), указ. изд.), у айнов (Horai et al., указ. изд.) и в Северо-Восточной Азии (Torroni et al., (1994), указ. изд.). Группа Е имеет свою собственную уникальную подгруппу на островах Юго-Восточной Азии, а также встречается на Тибете (типы Торрони 83, 89, 94, 104, 106, 109 и 119; последний идентифицирован неточно в связи с присутствием RFLP в точке 16517, Torroni et al. (1994), указ. изд.; илл. 2 в Metspalu et al., указ. изд.). Вновь описанный клад, М7 (возраст — 61 ... 20 тысяч лет), как и D, широко распространен в прибрежных районах Восточной Азии, но, подобно Е, вектор его распространения направлен в Юго-Восточную Азию: см. Kivisild et al. (2002), указ. изд.; Yong-Gang Yao, указ. изд. вернуться Данные для Центральной Азии — илл. 2 в Metspalu et al., указ. изд., для Тибета — Torroni et al., (1994), указ. изд., для Монголии — Kolman et al., указ. изд.; для Китая и Кореи — Torroni et al., (1994), указ. изд. вернуться Возраст четырех из этих ветвей в Монголии: Kivisild et al. (1999), указ. изд. (Обратите внимание, что существует возможность переоценки многообразия линий, влияющей на локальные показатели возраста этих линий в Монголии.) Средний возраст тех же линий на юге, в Южном Китае: табл. 3 в Yong-Gang Yao, указ. изд. Обратите внимание, что в Yong-Gang Yao, указ. изд., показано, что возраст индивидуальных ветвей D и G составляет 51—60 тысяч лет и что эти гаплогруппы совмещаются с М9/Е даже в более раннюю эпоху (Kivisild et al. (1999), указ. изд.). вернуться Группа В, возраст которой в Азии составляет около 75 тысяч лет (74 600 ... 18 700 лет, Yong-Gang Yao, указ. изд.), обладает наивысшей встречаемостью у коренных жителей Юго-Восточной Азии, Океании (за исключением горцев Новой Гвинеи и аборигенов Австралии) и западного побережья Тихого океана. |