Как видно, социальность и разум человека никак прямо не коррелированы с тотальной величиной его головного мозга.
Зато корреляция начинает проступать, когда измерению подвергаются по отдельности длиннотный, широтный и высотный диаметры роста головного мозга в филогенетической цепи: шимпанзе – австралопитек – археоантроп – палеоантроп – неоантроп. До неоантропов наиболее интенсивно увеличивался тотальный длиннотный размер (за счет роста задних областей); напротив, только мозг неоантропов дает интенсивный сдвиг высотного диаметра: в лобной и теменной долях эпицентры роста перемещаются из нижних отделов в верхние, происходит, как мы уже отмечали, усиленный рост префронтальной области, а также поднятие и выравнивание поверхности мозга в его своде.
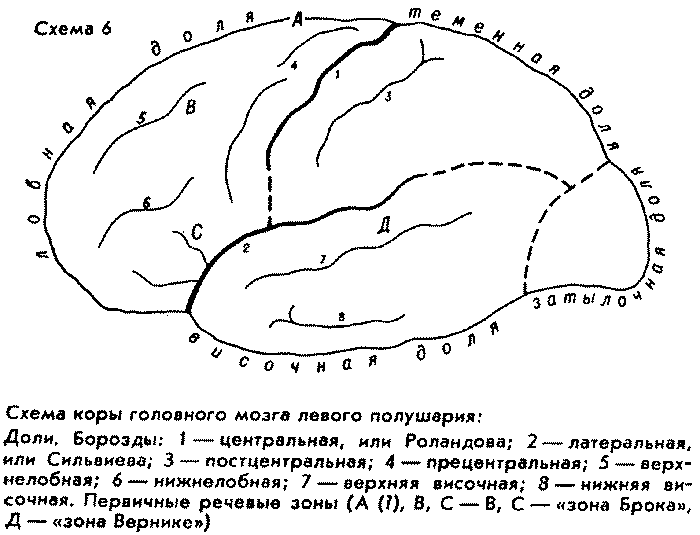
Интересно, что как раз в самой высотной, и тем самым весьма молодой, точке свода нашего мозга предположительно локализуется самый исходный, "инициальный" очажок речевой функции человека (см. схему коры мозга). Правда, вопрос пока является дискуссионным и открытым. Это так называемая "речевая зона Пенфильда", или "верхняя речевая кора", – якобы зона начальной речевой активизации. Эту зону Пенфильд и Роберте якобы обнаружили при оперативных исследованиях эпилепсии в так называемом дополнительном моторном поле на медиальной (внутренней, т.е. обращенной к другому полушарию) поверхности в задних отделах верхней лобной извилины. Некоторые авторы считают это прочно установленным фактом, другие при применении иных нейрохирургических методов не обнаруживают нарушений речевых функций при поражениях этой зоны (А. Р. Лурия) или замечают воздействие всего лишь на оттенки интонации и модуляции. Впрочем, последнее не служило бы решающим возражением: кто знает, может быть, так и рождалась первичная дифференциация звуков-знаков. Не будем пока ни принимать, ни отбрасывать гипотезу Пенфильда – Робертса. Заметим лишь, что, если бы указанная зона в самом деле все-таки оказалась начальным очажком второй сигнальной системы, было бы поразительно, что он расположен в самой высотной части, т.е. на самом молодом верхнем крае коры мозга Homo sapiens. В пользу второсигнальной принадлежности этого образования (если принять описание его Пенфильдом и Робертсом) говорит его односторонняя локализованность в доминантном полушарии (в левом у правшей), тогда как близко расположенные центры локализации находятся в обоих полушариях.
Интересно, что довольно высоко в структуре коры головного мозга Homo sapiens, впереди от средней части прецентральной извилины, лежит и участок, управляющий элементарными графическими действиями. Это свидетельствует, что они возникают на весьма ранних стадиях генезиса второй сигнальной системы. Люди начальной поры верхнего палеолита уже "рисовали", еще не вполне умея "разговаривать", если применять эти термины в современном смысле. Туда, в эту древность, уходят корни письменной речи. Зато когда иные археологи фантазируют, будто они обнаружили "рисунки" мустьерцев (палеоантропов, неандертальцев), мы уверенно можем считать это исключенным в той же мере, как заявку на изобретение перпетуум-мобиле: в мозге палеоантропов еще не было той высотной части, где находится центр, управляющий графическими действиями; они не могли носить и неуправляемого двигательно-подражательного характера (какие осуществляют шимпанзе), так как подражать в "рисовании" еще было тогда некому.
Еще далее вперед от самой высотной части свода головного мозга Homo sapiens (где сходятся лобная и теменная доли) лежит тот префронтальный отдел, о роли которого, в особенности его верхней формации, мы уже говорили в связи с постановкой проблемы происхождения второй сигнальной системы. Это образование присуще только Homo sapiens.
Можно сказать, что это главное морфологическое звено второй сигнальной системы. Отсюда по исключительно богатым нервным путям и контактам происходит возбуждение, активация (с помощью сетевидной, или ретикулярной, формации) отвечающих "задаче" или "намерению" двигательных центров мозга и одновременное торможение всех других двигательных центров, не идущих к делу.
Эволюционная морфология мозга не может ничего прямо сказать о развитии функциональной асимметрии полушарий (доминантность – субдоминантность, правшество – левшество). Вообще говоря, асимметрия как один из аспектов парности работы полушарий головного мозга обнаружена у животных: симметричные центры двух полушарий могут в данный момент находиться во взаимном антагонизме, т.е. в одном полушарии выполнять функцию возбуждения, во втором -торможения. Но у человека асимметрия закреплена: лишь кора одного полушария, обычно левого (у правшей), управляет всей второсигнальной функцией. Афазиологией доказана левосторонность управления динамикой как речевой, так и рече-мыслительной деятельности в лобных долях. Однако пока недостаточно подчеркивается, что вся эта односторонность управления второй сигнальной системой, т.е. локализация всех прямо причастных к ней зон и центров в "доминантном" полушарии, означает сопряженное торможение центров, управляющих неречевыми движениями – преимущественно с противоположного, "субдоминантного" полушария, но отчасти с того же. Если так, можно существенно переакцентировать обычное восприятие понятий "правшество", "левшество": дело не в том, что у "правши" правая рука (и другие органы) обладает некими повышенными возможностями, а в том, что у него, напротив, приторможена, снижена левосторонняя моторика (в первую очередь руки), у "левши" – наоборот. Есть немало оснований ограничить недавнее слишком абсолютное представление о двигательном "правшестве" и "левшестве": в большинстве случаев то и другое как – то смешано в индивиде. Но по некоторым данным онтогенеза можно предполагать, что в филогенезе асимметрия прошла три фазы: правшество – левшество – снова правшество; поэтому их следы у нас могут наслаиваться друг на друга. Что же мы обнаружили, выясняя древнейшие уровни, "исходные рубежи" второй сигнальной системы с помощью эволюционной морфологии мозга, афазиологии, генетической психологии? Мы обнаружили, что древнейшие зоны речевой деятельности возникают в моторной (двигательной), а не сенсорной (чувствующей) области коры. Это и отвечает выдвинутому выше тезису, что вторая сигнальная система родилась как система принуждения между индивидами: чего не делать; что делать. Мы получим дальнейшее подкрепление этого тезиса, перейдя от тотальных параметров, характеризующих особенности головного мозга Homo sapiens, к эволюционному сравнению состояния отдельных долей, извилин и борозд, поскольку они отразились на эндокранах ископаемых предковых форм, и к частным видам афазий.
Подчас антропологи принимают намеченное на эндокране питекантропа образование на месте так называемой "зоны Брока" в нижней лобной извилине за свидетельство присутствия у него речи, исходя из того, что у современного человека повреждения этой зоны вызывают нарушение двигательной (произносительной) речевой функции – моторную афазию. Со своей стороны повреждения "зоны Вернике" в первой височной извилине вызывают сенсорную афазию – нарушение восприятия чужой слышимой речи. Но речь не просто наличие двух устройств – передающего сигналы и принимающего их, тем менее – одного из двух; недостаточно и двусторонней связи между ними, т.е. единства слухо-двигательного анализатора. Допустим, питекантропы могли издавать и принимать разные крики; это не членораздельность, не речевые знаки в смысле современной науки. В. В. Бунак на основе анатомии периферических органов речи – гортани, челюстного скелета – установил, что у форм, предшествовавших Homo sapiens, не могло быть членораздельной синтагмической речи (дедукция какой-то якобы предшествовавшей, "нечленораздельной речи" не удалась – она противоречит всей аксиоматике науки о речи); в 1971 г. появилось сообщение, что Ф. Либерман (ун-т Коннектикут) изготовил искусственный макет гортани неандертальца (шапелльца) и получил экспериментальное физическое подтверждение того, что она не могла бы производить членораздельных звуков.